










JMLS 2022 December;7(2):163-170. 10.23005/ksmls.2022.7.2.163 Epub 2022 December 14
Copyright © 2022 by The Korean Society of Marine Life Science
Comparison of Biomarkers of Haliotis discus hannai and Hybrid Abalone in Marine Net Cage
Hyeon Jin Kim; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
So Ryung Shin; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
Seong Jin Kim; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
Jung Jun Park; Aquaculture Research Division, National Institute of Fisheries Science, Busan 46083, Korea
Jung Sick Lee; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
- Abstract
본 연구에서는 해상가두리 양식장에서 북방전복과 둥근전복속 교잡종(왕전복♀*둥근전복♂)의 생물지표를 분석하여 교잡육종의 결과를 평가하고자 하였다. 생존율은 북방전복과 유사하였으나, 성장(각장)은 교잡종에서 약 10% 빠른 것으로 분석되었다. 패각의 호흡공 기형율은 교잡종이 북방전복보다 약 6% 낮았으며, 패각 함몰 및 부식율은 교잡종이 약 15% 낮았다. 생화학적 조성에서는 조단백질의 경우 교잡종에서 약 3.1% 높았으며, 이를 제외한 나머지에서 유사한 값을 나타냈다. 소화, 흡수 및 해독기능을 수행하는 간췌장의 조직학적 평가에서는 교잡종에서 좋은 결과를 보였다. 이러한 결과로 보아 둥근전복속 교잡종은 추후 양식 환경에 접목하였을 때 높은 양식 생산력을 가질 것으로 판단된다.
In this study, the results of hybridization were evaluated by analyzing the biomarkers of Haliotis discus hannai and hybrid abalone (H. madaka♀*H. discus discus♂) in marine net cage. The survival rate was similar both experimental groups, but the growth (shell length) was about 10% faster in hybrid abalone. The deformity of respiratory pore in the hybrid abalone was about 6% lower than H. discus hannai, and the shell depression was about 15% lower in the hybrid abalone. In the biochemical composition, crude protein was about 3.1% higher in hybrid abalone, and showed similar values except for the crude protein. In the histological structure of the hepatopancreas, which performs the functions of digestion, absorption and detoxification of abalone, good results were obtained in the hybrid abalone. On these results, it is judged that the hybrid abalone will have high aquaculture productivity in the aquaculture environment.
Keywords: Hybrid abalone(교잡전복) Survival rate(생존율) Growth(성장) Shell deformity (패각기형) Hepatopancreas(간췌장)
Correspondence to: Jung Sick Lee; Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
- Received
- 15 November 2022;
- Revised
- 20 November 2022;
- Accepted
- 29 November 2022.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
전복류는 세계적인 고부가 산업 양식종으로 생산량이 지속적으로 증가하고 있는 추세이다(FAO, 2022). 한국의 전복 양식 생산량은 2000년에는 20톤이었으나 해상가두리 양식이 본격화되면서 2005년에는 2,062톤, 2010년에는 6,228톤, 2021년에는 23,199톤으로 생산량이 급격하게 증가하였다(KOSIS, 2022). 한국 해상가두리에서 주로 양식되는 종은 북방전복 Haliotis discus hannai과 둥근전복 H. discus discus인데 해상 양식장 지역 내 가두리 밀집, 고수온, 적조, 저질오염 등의 환경요인 악화에 따라 생산량 증가율이 감소하고 있다(Kim et al., 2018; Kim et al., 2021). 이러한 상황에서 생산량을 증가시키기 위한 육종기술 및 신품종 개발을 위한 다양한 연구들이 한국뿐만 아니라 미국, 중국, 일본, 호주 등에서 수행되면서 기존의 전복류보다 높은 생존율과 속성장, 환경적응력 등을 갖는 신품종들이 개발되고 있다(Hara, 1990; Hara and Kikuchi, 1992; Palada-de Vera and Eknath, 1993; Viana, 2002; Kube et al., 2007; Owen and Daniel, 2012; Kim et al., 2018; Kim et al., 2021; Ma et al., 2022).
전복류의 생물지표 개선을 위한 연구 결과, 중국에서는 H. discus hannai와 H. fulgens의 교잡종과 H. gigantea와 H. discus hannai의 교잡종을 개발하였으며(You et al., 2019; Boamah et al., 2020; Xiao et al., 2022; Ma et al., 2022), 칠레에서는 H. discus hannai와 H. rufescens의 교잡종 개발이 보고되었다(Lafarga-De la Cruz et al., 2013). 일본에서는 Nordotis gigantea와 N. discus (Miyaki et al., 1995), 멕시코에서는 H. rufescens와 H. corrugate (Tripp-Valdez et al., 2021), 호주에서는 H. rubra와 H. laevigata (Hamilton et al., 2009; Alter et al., 2017)의 교잡종을 개발하였다. 한국에서는 H. discus hannai와 H. discus discus (Kim et al., 2018; Park, 2020)의 종간 교잡을 통하여 새로운 품종을 개발한 사례가 있다. 하지만 이들의 양식산업 적용을 위해서는 해당 교잡종에 대한 양식 현장에서의 생물지표를 평가할 필요가 있다. 따라서 본 연구에서는 둥근전복속 교잡종(왕전복 H. madaka♀*둥근전복 H. discus discus♂)을 북방전복과 비교하여 양식 현장에서의 생존율 및 성장 효과 등을 알아보고자 하였다.
재료 및 방법
1. 재료
연구에는 인공수정을 통해 생산된 둥근전복속 교잡종(왕전복 H. madaka♀*둥근전복 H. discus discus♂)을 이용하였으며, 비교종으로 북방전복 H. discus hannai를 사용하였다. 인공수정에 사용된 모패는 왕전복과 둥근전복은 각각 강원도 고성의 저도 인근 해역과 제주도 인근 해역에서 다이버를 투입하여 채집하였다. 실험에는 외형과 부착능력에 이상이 없는 개체로 평균 각장 24.5±4.0 mm, 전중 1.3±0.5 g의 북방전복과 평균 각장 22.2±2.8 mm, 전중 1.2±0.4 g 크기의 교잡종을 사용하였다.
2. 방법
1) 사육조건
본 연구는 전남 완도군 보길면에 위치한 해상 가두리 양식시설(2.4 m×2.4 m×3 m)에 각각 750개체를 수용하여 36주 동안 2회 반복구를 이용하여 수행하였다(Fig. 1).

2) 생물지표 분석
(1) 생존율
실험생물의 사망은 육질부가 없는 개체의 누적사망률을 구하여 생존율로 환산하여 나타냈다.
(2) 계측형질
시료의 각장과 각폭은 쉘터에 부착한 측정용 자를 실험생물과 함께 촬영한 후, 화상분석 프로그램(i-solution, IMT Inc., U.S.A.)을 이용하여 측정하였으며, 전중과 육중량은 전자저울을 이용하여 측정하였다(Fig. 2).

(3) 비만도(condition index, CI)
비만도는 다음과 같은 식으로 계산하였다.
▣ Condition index = Body weight (g)/Shell length (mm)3 × 100
(4) 패각 기형률
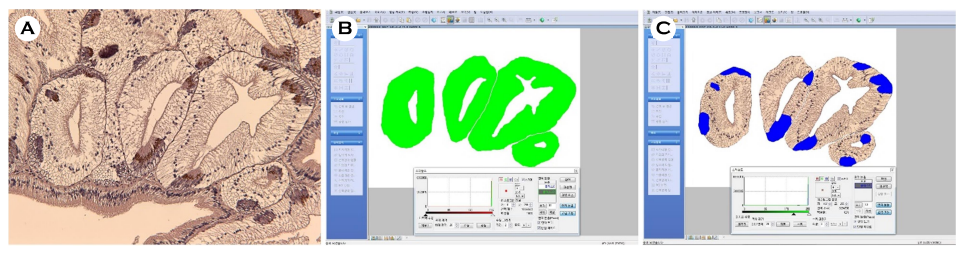
패각 기형의 형태는 호흡공 기형(Fig. 3A and B)과 패각의 함몰 및 부식(Fig. 3C) 두 가지로 구분하여 각각의 백분율(%)로 나타냈다.

(5) 생화학적 조성
일반성분은 AOAC (Association of official analytical chemist, U.S.A.)법으로 분석하였으며, 수분함량은 105℃ 건조법, 회분은 550℃ 회화법으로 분석하였다. 조지방 분석은 조지방추출기(SER 158/6, VELP Scientifica Sri, Italy)를 이용하였으며, 조단백질은 원소분석기(vario MACRO cube, DKSH Ltd., New Zealand)를 이용하여 전질소량을 정량하고 질소계수 6.25를 곱하여 나타냈다. 탄수화물은 100에서 수분, 조단백질, 조지방, 회분의 값을 제외한 값으로 하였다.
3) 광학현미경 표본제작
광학현미경 표본은 시료를 해부하여 간췌장의 일부를 10% 중성포르말린에 24시간 동안 고정하였다. 고정된 시료는 흐르는 물에 수세한 후 70~100% 에탄올로 단계별 탈수를 진행하였다. 파라핀 포매 후 마이크로톰(RM2235, Leica, Germany)을 이용하여 4~6 μm 두께로 연속절편을 제작하여 Mayer's hematoxylin-eosin (H-E) 염색 후 광학현미경으로 분석하였다.
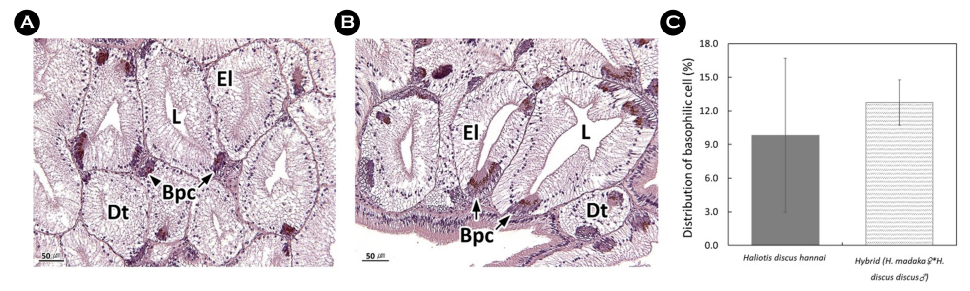
4) 호염기성세포 분포율
소화선세관을 구성하는 호염기성세포의 분포율을 확인하기 위해 광학현미경 표본을 JPEG 파일로 이미지화 하였다. 호염기성세포의 분포율은 화상분석 프로그램(i-solution, IMT Inc., U.S.A.)를 이용하여 아래의 식을 통해 백분율(%)로 나타냈다(Fig. 4).
Distribution of basophilic cell (%) = Basophilic cell area (μm2)/Epithelial layer area of digestive tubules (μm2) × 100
결 과
1. 생존율
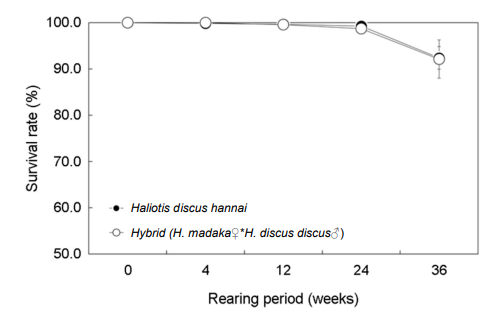
사육 4주 후 생존율은 북방전복은 99.9%, 교잡종은 100%였으며, 12주 후에는 각각 99.7%와 99.6%, 24주 후에는 각각 99.3%, 98.7%였으며, 36주 후에는 북방전복 92.4%, 교잡종 92.1%로 유사하였다(Fig. 5).
2. 성장
1) 각장
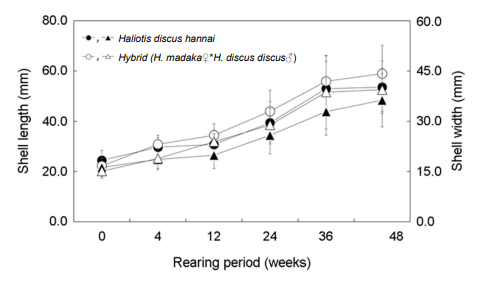
해상가두리 사육 개시기에 평균 각장은 북방전복 24.5±4.0 mm, 교잡종 21.0±3.6 mm였다. 4주 후에는 각각 29.7 mm와 30.8 mm, 12주 후에는 각각 30.8 mm와 34.4 mm, 24주 후에는 북방전복 39.4 mm, 교잡종은 44.0 mm였다. 36주 후 평균 각장은 북방전복 53.0±10.9 mm, 교잡종 55.9±10.1 mm로 북방전복에 비해 교잡종의 성장이 약 10.0% 빠른 것으로 나타났다(Fig. 6).
2) 각폭
평균 각폭은 개시기에 북방전복 16.2 mm, 교잡종 15.1 mm였다. 36주 후 평균 각폭은 북방전복 32.9±7.1 mm, 교잡종 38.7±11.0 mm로 북방전복과 비교하였을 때 교잡종이 약 8.6% 빠른 각폭 성장을 보였다(Fig. 6).
3) 전중
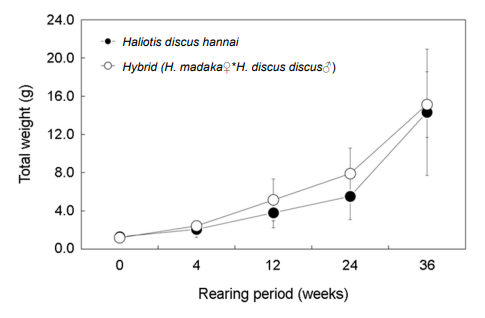
개시기의 전중은 북방전복과 교잡종 각각 1.3 g, 1.2 g이었으며, 12주 후에는 각각 3.8 g과 5.1 g이었으며, 36주 후에는 북방전복 14.3 g, 교잡종 15.1 g으로 교잡종이 약 5.6% 빠른 성장을 보였다(Fig. 7).
4) 육중량
육중량은 개시기에 북방전복 0.9 g, 둥근전복속 교잡종 0.7 g이었으며, 12주 후에는 각각 2.6 g과 3.6 g, 24주 후에는 각각 3.8 g과 5.4 g으로 나타났다. 36주 후의 육중량은 북방전복과 둥근전복속 교잡종 각각 8.9 g, 9.6 g으로 교잡종이 약 7.4% 빠른 성장을 보였다(Fig. 8).
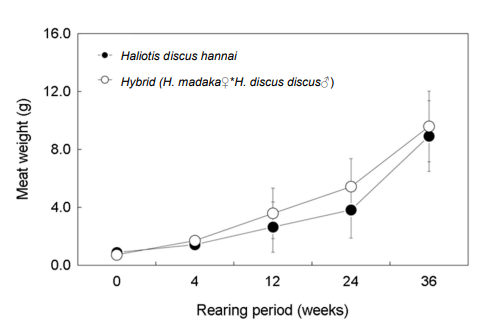
Fig. 8. Meat weight of Haliotis discus hannai and hybrid abalone (H. madaka♀*H. discus discus♂) for 36 weeks in marine net cage. Vertical bar: SD.
5) 비만도
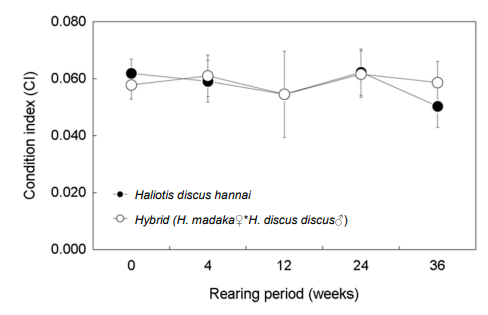
비만도는 개시기에 북방전복은 0.062, 둥근전복속 교잡종은 0.058를 나타냈다. 36주 후에는 북방전복과 교잡종 각각 0.050과 0.059로 북방전복에 비해 교잡종이 높았다(Fig. 9).
3. 패각 기형률
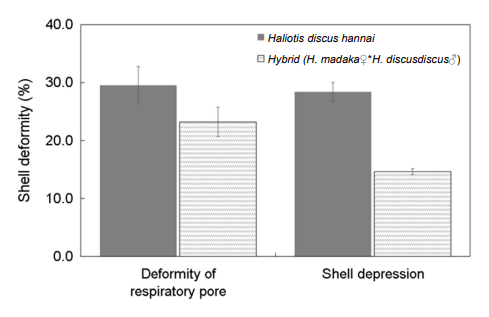
패각 기형 가운데 호흡공 기형은 북방전복 29.5%, 교잡종 23.2%였다. 패각 함몰 및 부식은 북방전복 29.5%, 교잡종 14.6%로 북방전복에 비해 교잡종에서 약 15% 낮았다(Fig. 10).
4. 생화학적 조성
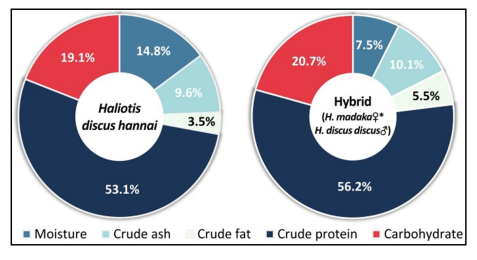
생화학적 조성 가운데 탄수화물은 북방전복 19.1%, 교잡종 20.7%였으며, 조회분은 9.6%, 10.1%로 유사하였다. 조지방은 각각 3.5%와 5.5%였으며, 조단백의 구성비는 교잡종에서 56.2%로 북방전복의 53.1%에 비해 높았다(Fig. 11).
5. 간췌장 건강성
간췌장은 다수의 소화선세관으로 이루어져 있으며, 소화선세관은 단층 상피층으로 상피세포와 H-E 염색 표본에서 세포질이 호염기성을 나타내는 호염기성세포로 구성된다. 실험구별 간췌장의 조직학적 분석 결과, 북방전복에서는 일부 상피세포의 변성과 호염기성세포의 위축이 관찰되었으나, 교잡종에서는 북방전복에 비해 상피세포의 변성 정도가 낮았다(Fig. 12A and B). 호염기성세포의 분포율은 북방전복과 교잡종에서 각각 9.8%와 12.8%로 교잡종에서 높은 값을 보였다(Fig. 12C).
고 찰
전복류의 종간 교배는 전복 양식에서의 생산량 증가 전략 중 하나로 생존율, 성장률, 현장 적응력, 전복의 품질 향상에 긍정적인 영향을 미친다(Robinson et al., 2010; Lafarga-De la Cruz and Gallardo-Escárate, 2011).
전복의 종에 따른 생존율 차이는 여름철에 교잡종(H. discus hannai♀*H. fulgens♂, H. fulgens♀*H. discus hannai♂)이 폐사율 5% 미만으로 북방전복 H. discus hannai에 비해 상대적으로 높은 생존율을 보였으며(You et al., 2015), 둥근전복속 교잡종(H. discus hannai♀*H. discus discus♂) 역시 북방전복에 비해 생존율이 높았다(Kim et al., 2018; Park, 2020). Boamah et al. (2020)은 염분 농도 28, 30, 32 및 34 psu에서 모두 두 교잡종(H. gigantea♀*H. discus hannai♂, H. discus hannai♀*H. fulgens♂)이 북방전복에 비해 높은 생존율을 보고하였다. H. rufescens♀와 H. corrugata♂의 교잡종은 수온 22℃에서 H. rufescens에 비해 약 23% 높은 생존율을 보였다(Tripp-Valdez et al., 2021). Xiao et al. (2022)에 따르면 고수온 시기(7~9월)에 교잡종(H. gigantea♀*H. discus hannai♂)이 H. gigantea와 H. discus hannai에 비해 생존율이 높았다. 그러나, 교잡전복(N. gigantea♀*N. discus♂)에서는 N. gigantea에 비해 낮은 생존율을 보였으며(Miyaki et al., 1995), 부화 461일 후 교잡종(H. rufescens♀*H. discus hannai♂)의 생존율은 H. discus hannai보다 높았으나, H. rufescens에 비해 낮은 값을 보였다(Lafarga-De la Cruz and Gallardo-Escárate, 2011). 본 연구에서는 둥근전복속 교잡종과 북방전복의 생존율은 유사하였다.
교잡전복의 성장은 여름철에 교잡전복(H. discus hannai♀*H. fulgens♂)이 순종보다 약 10.4~16.1% 빠른 각장 성장을 보였으며(You et al., 2015), 교잡전복(H. gigantea♀*H. discus hannai♂)에서도 순종에 비해 각장과 전중 성장이 유의적으로 높았다(Xiao et al., 2022). 또한, 모든 염분 조건에서 교잡전복(H. gigantea♀*H. discus hannai♂, H. discus hannai♀*H. fulgens♂)의 전중, 체중의 성장률이 북방전복 H. discus hannai보다 높았다(Boamah et al., 2020). 그러나, 교잡전복(N. gigantea♀*N. discus♂)은 4~9월까지는 순종에 비해 느린 각장 성장을 보였으나, 10월부터는 순종보다 성장률이 높았다(Miyaki et al., 1995). 본 연구는 교잡종이 북방전복에 비해 빠른 성장을 보이고 있어 이들 연구와 유사한 결과를 보였다.
복족류의 패각 기형은 생물학적인 관점에서 중요할 뿐만 아니라 잠재적인 환경오염 지표로 사용될 수 있다(Zuykov et al., 2012). 복족류와 이매패류의 패각은 외투막의 분비세포들에 의해서 형성되며, 다양한 화학적 오염원에 의해 외투막은 구조 및 기능적 저하를 나타낸다(Park et al., 2011; Kim et al., 2021). 본 연구에서는 교잡종이 북방전복에 비해 패각 기형률이 낮았는데, 이는 수중의 환경 오염물질, 고수온 등에 대한 높은 환경적응력에 의한 것으로 판단된다.
전복을 포함한 패류의 생화학적 조성은 먹이, 성장, 계절, 해양 산성화 및 환경 변화에 의해 영향을 받는다(Allen et al., 2006; Mateose et al., 2010; Ju et al., 2015; Anacleto et al., 2014; Tate et al., 2017; Boamah et al., 2020). 생화학적 조성 중 단백질은 주요 체구성 성분으로 장기간의 스트레스 상황에서는 대사과정의 에너지원으로 사용된다(Lemasson et al., 2018). 교잡전복(H. gigantea♀*H. discus hannai♂, H. discus hannai♀*H. fulgens♂)과 순종 전복의 일반성분 분석에서 조단백질의 유의적인 차이는 관찰되지 않았다(Boamah et al., 2020). 본 연구에서는 교잡종이 북방전복에 비해 조단백질의 함량이 높게 나타나 환경적응력이 높을 것으로 판단된다.
복족류의 간췌장은 소화, 저장 및 해독기능을 수행하는 기관으로 다수의 소화선세관으로 구성되며, 소화선세관은 단층 상피층으로 상피세포와 호염기성세포로 구분된다(Zaldibar et al., 2007; Jung et al., 2013; Kim et al., 2021). 본 연구에서 간췌장의 건강성을 조직학적으로 분석한 결과, 교잡종에서는 북방전복에 비해 상피세포의 변성 정도가 낮고, 호염기성세포의 분포비율이 높아 소화, 흡수 및 해독기능의 정상적인 수행이 가능한 것으로 보아 해상가두리에서의 높은 적응력을 가질 것으로 판단된다.
이와 같은 결과로 보아 둥근전복속 교잡종(왕전복 H. madaka♀*둥근전복 H. discus discus♂)은 해상가두리에서 높은 적응력을 가지며, 양식 산업의 적용을 고려할 만하다고 판단되지만, 추후 장기간 사육에 따른 생물지표 변화를 확인할 필요가 있다.
- References
-
1. Allen JV, Marsden DI, Ragg LCN, Gieseg S. 2006. Effects of tactile stimulants on feeding, growth, behavior, and meat quality of cultured Blackfoot abalone, Haliotis iris. Aquaculture 257: 294-304.
-
2. Alter K, Andrewartha SJ, Morash AJ, Clark TD, Hellicar AD, León RI, Elliott NG. 2017. Hybrid abalone are more robust to multi-stressor environments than pure parental species. Aquaculture 478: 25-34.
-
3. Anacleto P, Maulvault AL, Bandarra NM, Repolho T, Nunes ML, Rosa R, Marques A. 2014. Effect of warming on protein, glycogen and fatty acid content of native and invasive clams. Food Res Int 64: 439-445.
-
4. Boamah GA, Wang T, Chowdhury IA, Luo X, Huang M, Xu C, Ke C, You W. 2020. Sub-low salinity impact on survival, growth and meat quality of the pacific abalone (Haliotis discus hannai) and hybrids. Aquac Res 51: 5184-5193.
-
5. Food and Agriculture Organization of the United Nations (FAO). 2022. FishStatJ-Software for Fishery and Aquaculture Statis- tical Time Series. Statistics Team (NFISS) Fisheries and Aqua- culture Division (NFI). Food and Agriculture Organization of the United Nations, Rome, Italy. https://www.fao.org/fishery /statistics/software/fish
-
6. Hamilton MG, Kube PD, Elliott NG, McPherson LJ, Krsinich A. 2009. Development of a breeding strategy for hybrid abalone. Proc Assoc Advmt Anim Breed Genet 18: 350-353.
-
7. Hara M, Kikuchi S. 1992. Increasing growth rate of abalone Haliotis discus hannai, using selective techniques. NOAA Tech Rep NMFS 106: 21-26.
-
8. Hara M. 1990. The effect of genetics of growth in three groups of abalone seed. Bull Tohoku Natl Fish Res Inst 52: 73-78.
-
9. Ju ZY, Viljeon C, Hutchinson P, Reinicke J, Horgen DF, Howard L, Lee CS. 2015. Effects of diets on the growth performance and shell pigmentation of Pacific abalone. Aquac Res 47: 1-11.
-
10. Jung GK, Park JJ, Jeon MA, Shin HC, Lee YG, Lee JS. 2013. Hepato- pancreas ultrastructure of the spiny top shell, Batillus cornutus (Gastropoda: Turbinidae). Bull Fish Sci Inst Chonnam Nat Univ 21: 11-15.
-
11. Kim HJ, Shin SR, Lee JS, Hwang D, Kim JW. 2021. Biomarkers with dietary conditions of juvenile hybrid abalone (Haliotis discus discus♀*H. madaka♂). J Korean Soc Fish Ocean Technol 57: 365-373.
-
12. Kim J, Kim S, Lee JS, Kim JW. 2018. Tolerance on high water temperature of hybrid abalone (Haliotis discus hannai♀*H. discus discus♂). J Kor Soc Fish Mar Edu 30: 2102-2111.
-
13. Korean Statistical Information Service (KOSIS). 2022. Domestic Statistics. Korean Statistical Information System. https:// kosis.kr/index/index.do
-
14. Kube PD, Appleyard SA, Elliott NG. 2007. Selective breeding greenlip abalone (Haliotis laevigata): preliminary results and issues. J Shellfish Res 26: 821-824.
-
15. Lafarga-De la Cruz F, Gallardo-Escárate C. 2011. Intraspecies and interspecies hybrids in Haliotis: natural and experimental evidence and its impact on abalone aquaculture. Rev Aquac 3: 74-99.
-
16. Lafarga-De la Cruz F, Núñez-Acuña G, Gallardo-Escárate C. 2013. Hybridization between Haliotis rufescens and Haliotis discus hannai: evaluation of fertilization, larval development, growth and thermal tolerance. Aquac Res 44: 1206-1220.
-
17. Lemasson AJ, Hall-Spencer JM, Kuri V, Knight AM. 2018. Changes in biochemical and nutrient composition of seafood due to ocean acidification and warming. Mar Environ Res 143: 82-92.
-
18. Ma YB, Zou WG, Ai CX, You WW, Liu ST, Luo X, Ke CH. 2022. Evaluation of optimal dietary protein levels for juvenile hybrid abalone under three temperatures: growth performance, body composition, biochemical responses, and antioxidant capacity. Aquac Nutr 2022: 7008746.
-
19. Mateos HT, Lewandowski PA, Su XQ. 2010. Seasonal variations of total lipid and fatty acid contents in muscle, gonad, and digestive glands of farmed Jade Tiger hybrid abalone in Australia. Food Chem 123: 436-441.
-
20. Miyaki K, Niiyama H, Tabeta O. 1995. Growth and survival of arti- ficial hybrid abalone, Nordotis gigantea female × N. discus male under laboratory rearing conditions. Bull Jpn Soc Sci Fish 43: 401-405.
-
-
22. Palada-de Vera MS, Eknath AE. 1993. Predictability of individual growth rates in tilapia. Aquaculture 111: 147-158.
-
23. Park H. 2020. Physiological responses of Pacific abalone (Haliotis discus hannai) and hybrid abalone (Haliotis discus hannai × Haliotis discus discus) exposed to temperature and ammonia. Doctor's Thesis, Pukyong National University, Busan.
-
24. Park JJ, Kim S, Park MA, Lee JS. 2011. Ultrastructural changes in the mantle of the equilateral venus, Gomphina veneriformis (Bivalve: Veneridae) exposed to TBTCl. Korean J Malacol 27: 213-221.
-
25. Robinson N, Li X, Hayes B. 2010. Testing options for the commer- cialization of abalone selective breeding using bioeconomic simulation modelling. Aquac Res 41: 268-288.
-
26. Tate RD, Benkendorff K, Ab Lah R, Kelaher BP. 2017. Ocean acidifi- cation and warming impacts the nutritional properties of the predatory whelk, Dicathais orbita. J Exp Mar Biol Ecol 493: 7-13.
-
27. Tripp-Valdez MA, Cicala F, Galindo-Sánchez CE, Chacón-Ponce KD, López-Landavery E, Díaz F, Re-Araujo D. 2021. Growth performance and transcriptomic response of warm-acclimated hybrid abalone Haliotis rufescens (♀) × H. corrugata (♂). Mar Biotechnol 23: 62-76.
-
-
29. Xiao Q, Shen Y, Gan Y, Wang Y, Zhang J, Huang Z, You W, Luo X, Ke C. 2022. Three-way cross hybrid abalone exhibit het- erosis in growth performance, thermal tolerance, and hypoxia tolerance. Aquaculture 555: 738231.
-
30. You W, Guo Q, Fan F, Ren P, Luo X, Ke C. 2015. Experimental hybridization and genetic identification of Pacific abalone Haliotis discus hannai and green abalone H. fulgens. Aqua- culture 448: 243-249.
-
31. You W, Wang B, Luo X, Ke C. 2019. Environmental stress tolerance and immune response for the small abalone hybrids. Aquac Int 27: 105-123.
-
32. Zaldibar B, Cancio I, Marigómez I. 2007. Reversible alterations in epithelial cell turnover in digestive gland of winkles (Littorina littorea) exposed to cadmium and their implications for bio- marker measurements. Aquat Toxicol 81: 183-196.
-
33. Zuykov M, Vinarski M, Pelletier E, Demers S, Harper DA. 2012. Shell malformations in seven species of pond snail (Gastropoda, Lymnaeidae): analysis of large museum collections. Zoosyst Evol 88: 365-368.
Copyright (C) 2024. The Korean Society of Marine Life Science