










JMLS 2022 December;7(2):113-120. 10.23005/ksmls.2022.7.2.113 Epub 2022 December 14
Copyright © 2022 by The Korean Society of Marine Life Science
Exposure to Sublethal Concentrations of Copper Pyrithione Reduces Cholinergic Activity and Induces Oxidative Stress in a Marine Polychaete
Md. Niamul Haque; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea; Research Institute of Basic Sciences, Incheon National University, Incheon 22012, Korea
Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea; Research Institute of Basic Sciences, Incheon National University, Incheon 22012, Korea; Yellow Sea Research Institute, Incheon 22012, Korea
- Abstract
Despite concerns about the significant toxicity of copper pyrithione (CuPT) at environmental concentrations, effects of CuPT on benthic organisms have received little attention. Here, we analyzed the detrimental effects of CuPT at sublethal concentrations (1/50, 1/20, and 1/10 of the 96 h-LC50 value) for 14 days in the marine polychaete Perinereis aibuhitensis. Reduced burrowing activity and significantly decreased the acetylcholinesterase activity in response to relatively high concentrations of CuPT were identified as CuPT-triggered cholinergic inhibition. The lipid peroxidation marker, malondialdehyde levels were dose-dependently increased, whereas intracellular glutathione was depleted by relatively high concentrations. In the CuPT-treated polychaete, significant fluctuations in the enzymatic activities of the antioxidant defense system (catalase, superoxide dismutase, glutathione reductase, and glutathione peroxidase) were observed with significantly modulated glutathione S-transferase activity. These results indicate that even sublethal levels of CuPT would have detrimental effects on the health status of the marine polychaete.
Keywords: Marine polychaete Copper pyrithione Acetylcholinesterase Lipid peroxidation Antioxidant defense Oxidative stress
Correspondence to: Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
- Received
- 6 September 2022;
- Revised
- 3 October 2022;
- Accepted
- 12 October 2022.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

Introduction
Copper pyrithione (CuPT) has been efficiently used as a sub- stitute of the biofoulant tributyltin in antifouling paints (Yebra et al., 2004). Antifoulants containing CuPT are an important source of elevated levels of Cu in marine environments, including in coastal sediments (Schiff et al., 2007). In addition, pyrithions (PTs) are widely used as fungicides, bactericides, or algicides in various commercial products such as agricultural insecticides, cosmetics, industrial fluids, coatings, and aquaculture tanks (Doose et al., 2004; Guthery et al., 2005). PTs affect the homeostasis of different metal cations as ionophores and they have induced teratogenic impacts on aquatic organisms such as sea urchins and fish at relatively low concentrations (< 20 ng l-1) (Goka, 1999; Thomas, 1999; Kobayashi and Okamura, 2002). Consequently, the incorporation of CuPT in antifoulants and its extensive use is expected to increase its con- centration in the aquatic environment. In laboratory and natural conditions, ZnPT can easily be transformed into CuPT by trans- chelation with Cu (Thomas, 1999; Grunnet and Dahllof, 2005). Despite its enormous application, there is still limited research on the toxicity of CuPT in marine invertebrates. Thus, it is essential to understand the harmful impacts of CuPT on non-target aquatic organisms.
It was reported that CuPT causes skeletal deformity via acetyl- cholinesterase (AChE) inhibition in Fundulus heteroclitus (Mochida et al., 2008). In the toxicological study, changes in AChE activity have been widely used as a promising indicator of neural toxicity (Nunes, 2011). Lipid peroxidation, damage of polyunsaturated fatty acids in the cell membrane by reactive oxygen species (ROS), is a chain reaction that produces malondialdehyde (MDA) (Lushchak, 2011). MDA is a toxic end-product of lipid peroxidation, which has been extensively used as an indicator of oxidative stress induction (Lushchak, 2011). The protective machinery in aquatic organisms comprises different antioxidant components, which include gluta- thione (GSH), superoxide dismutase (SOD), catalase (CAT), gluta- thione reductase (GR), glutathione peroxidase (GPx), and gluta- thione S-transferase (GST) (Lushchak, 2011). These antioxidant enzymes are involved in the biotransformation and detoxification of intracellular ROS and free radicals, which are generally increased during the oxidative stress conditions (Livingstone, 2001). Therefore, in this study, alterations in the neurotoxicity indicator AChE, GSH level, and antioxidant enzymes (SOD, CAT, GR, GPx, and GST) were employed to estimate the impacts of CuPT using the marine polychaete.
Due to small size, deposit feeders, ease of maintenance in laboratory, endurance in adverse environmental circumstances, sensitivity to various contaminants, and worldwide distribution, several polychaetes (e.g., Hediste species, Perinereis species) have been widely used in toxicological studies of benthic environment (Reish and Gerlinger, 1997; Dean, 2008; Rhee et al., 2012). The pre- sent study evaluated the detrimental effects of CuPT at sublethal concentrations through measurements of burrowing ability and AChE activity, following assessments of biochemical modulation of MDA, GSH, and antioxidant defense enzymes (i.e., SOD, CAT, GR, GPx, and GST) in the marine polychaete Perinereis aibuhitensis for 14 days. These results will help to determine whether CuPT can be employed as a permanent sustainable alternative of TBT and other toxic antifoulants.
Materials and Methods
1. Marine polychaete
The marine polychaete P. aibuhitensis was obtained from the polychaete aquaculture company in Yeosu, Jeollanam-do, South Korea. Approximately 5,000 polychaetes (≈ 1.62 ± 0.35 g; 3 months after fertilization; female:male ratio 6:4) were transferred and cul- tured under conditions of constant photoperiod (16 h:8 h L:D), 16 ± 0.8℃, 32 practical salinity unit, 7.5 ± 0.4 mg l-l DO, and pH 8.0 in opaque round polyethylene tanks with filtered seawater and sieved sediment (mixture of sand and mud as 8:2 ratio provided from the polychaetes aquaculture company) at Incheon National University, Incheon, South Korea. Fifty percent of the overlying filtered seawater was replaced every 2 days. The marine poly- chaetes were fed on ground TetraMarin® (Tetra, Blacksburg, VA, USA; 0.1 g per polychaete) with a mixture of microalgae (1 × 106 cells ml-1 containing Dunaliella sp., Isochrysis sp., and Tetraselmis sp.) in a ratio of cell number, 1:1:1) three times in a week. Salinity, temperature, DO, and pH were monitored twice daily using a portable Orion Star meter (Thermo Fisher Scientific, Tewksbury, MA, USA).
2. Exposure
CuPT [C10H10CuN2O2S2, 1-hydroxy-2(1h)-pyridinethione, copper] was purchased from ACROS OrganicsTM (CA, USA) and stock solution was prepared by dissolving in dimethyl sulfoxide (DMSO; Sigma-Aldrich Co, St. Louis, MO, USA). Designated nominal con- centrations of CuPT were prepared by serial dilution of the stock solution in filtered seawater. The final DMSO concentrations ranged from 0.01~0.1% in exposure volumes. The 0.1% DMSO showed no significant change in our previous study (Eom et al., 2019).
To measure the LC50 value, thirty polychaetes (n=10 per each concentration as triplicate) were treated with different concen- trations of CuPT in 55 × 45 × 36 cm opaque glass fiber tank (21st-century HighTech®, Busan, South Korea) in the absence of sediment. Exposure was kept under the same conditions as that of polychaete culture with trickle flow aeration. Half of the test solution was renewed daily with the addition of an equivalent concentration of CuPT. In the case of the control group, half of the filtered seawater was renewed daily. Determination of acute toxicity value (e.g., LC50) was conducted by Probit-analysis using ToxRat® Professional 2.10.3.1 (ToxRat Solutions GmbH, Alsdorf, Germany) at 96 h after exposure to waterborne CuPT. No food was supplied during the toxicity experiment. Dead polychaetes were immediately discarded from the test chamber. A preliminary range-finding experiment was conducted with different concen- trations of CuPT (i.e., 1, 10, 100, 500, 1,000, 1,500, and 2,000 μg l-1). In the definitive test, the following concentrations of CuPT were used: 0.1, 1, 5, 10, 50, 100, 200, 400, 600, 800, and 1,000 μg l-1.
To evaluate the molecular and biochemical responses of the marine polychaete upon sublethal concentrations, approximately 300 polychaetes were collected and 20 polychaetes were exposed to each concentration of CuPT (i.e., control, DMSO, 1/50, 1/20, and 1/10 of 96 h-LC50 value) in an opaque glass fiber tank (21st-century HighTech®) with 9~10 cm of sediment. The polychaetes were sampled at 0 h, 24 h, 96 h, day 7, and day 14 days. Cultures were kept under the same conditions of acclimation. Of 20 poly- chaetes, 15 polychaetes from each concentration were collected and divided into three sets as triplicates, and the three sets of pooled samples were used for the analysis of biochemical re- sponses. Half of the exposed solution was refreshed every 24 h with the addition of an equivalent concentration of CuPT.
3. Burrowing and AChE activity
Survived polychaetes were collected from day 14 on each con- centration of CuPT and were tested for their burrowing ability. Overall methods were followed by previous methodology on polychaete burrowing assays (Bonnard et al., 2009; Fonseca et al., 2018). Twenty survived polychaetes from each treatment were moved into clean filtered seawater for burrowing assay in 200 ml glass fiber container filled with sieved sediment (depth 5 cm). The number of fully burrowed polychaetes was counted every two minutes for 30 min.
AChE activity was determined in the CuPT-exposed polychaete by the Ellman method (Ellman et al., 1961). The pooled polychaete sample was homogenized with a Teflon homogenizer in 0.1 M phosphate buffer (pH 8.0) and centrifuged at 3,000 g for 30 min at 4℃. The separated supernatant was mixed with 0.1 M phos- phate buffer (pH 8.0) containing 5,5'-dithiobis (2-nitrobenzoic acid) (DTNB, 0.01 M; Sigma-Aldrich, Inc., St. Louis, MO, USA) and acetylthiocholine iodide (ATCh, 0.075 M; Sigma-Aldrich, Inc.) in a 3 ml cuvette. A blank without ATCh and a blank without sample were prepared and tested at 25℃ for 5 min to measure total AChE. The measurements were performed using a ThermoTM Varioskan Flash spectrophotometer (Thermo Fisher Scientific, Tewksbury, MA, USA) at an absorbance of 412 nm. The activity of AChE was normalized to total protein in the supernatant. Protein levels were determined following the method described by Bradford (1976).
4. Measurement of antioxidant parameters
Detailed descriptions for the measurement of MDA and anti- oxidant parameters are omitted as described in our previous study conducted with P. aibuhitensis (Rhee et al., 2012; Eom et al., 2019; Haque et al., 2020). GSH concentration was measured in the pooled polychaete sample after washing in 0.9% NaCl using GSH Assay Kit (Catalog No. CS0260; Sigma-Aldrich, Inc.). Enzymatic activities of CAT and SOD were analyzed by CAT Assay Kits (Catalog No. CAT100; Sigma-Aldrich, Inc.) and SOD Assay Kits (Catalog No. 19160; Sigma-Aldrich Chemie, Switzerland), respect- ively, following the manufacturer's instructions. Enzymatic activities of GR and GPx were analyzed by GR Assay Kit (Catalog No. GRSA; Sigma-Aldrich, Inc.) and GPx Assay Kit (Catalog No. CGP1; Sigma-Aldrich, Inc.), respectively, following the manufacturer's instructions. The GST activity was evaluated according to our previous studies on marine polychaete (Rhee et al., 2007).
5. Statistic analysis
Statistical data analysis was conducted with the statistical soft- ware package SPSS, Version 17 (SPSS Inc., Chicago IL, USA). A one-way analysis of variance (ANOVA) followed by a Duncan's multiple range test was used to evaluate significant differences between the control and treated groups. The data are expressed as means ± S.D. (standard deviation) and p value less than 0.05 was considered significant difference.
Results
1. Effects on the burrowing ability and AChE activity
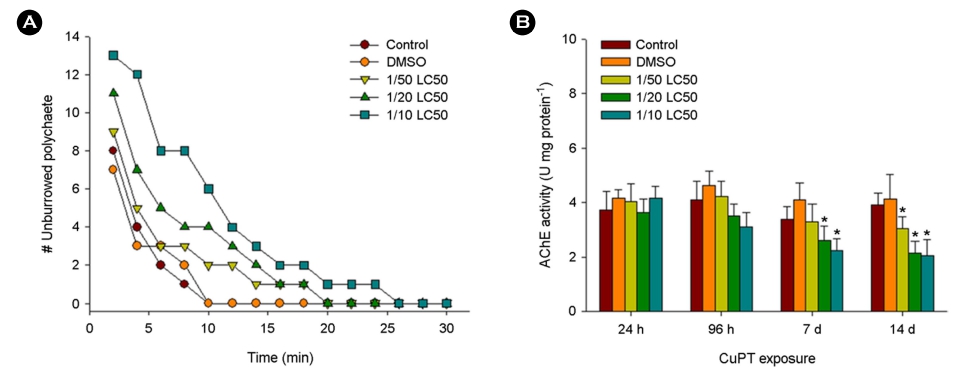
The 96 h-LC50 value upon CuPT treatment was 4.2 μg l-1. The number of unburrowed polychaetes was dose-dependently in- creased for 30 min (Fig. 1A). The burrowing activity of polychaetes was substantially delayed at the highest concentration of CuPT (1/10 LC50). A significant inhibition in AChE activity was observed after exposure to CuPT at 1/20 and 1/10 LC50 values on day 7 and 1/50, 1/20, and 1/10 LC50 values on day 14 compared to the control (p < 0.05) (Fig. 1B).
2. Effects on the MDA and GSH contents
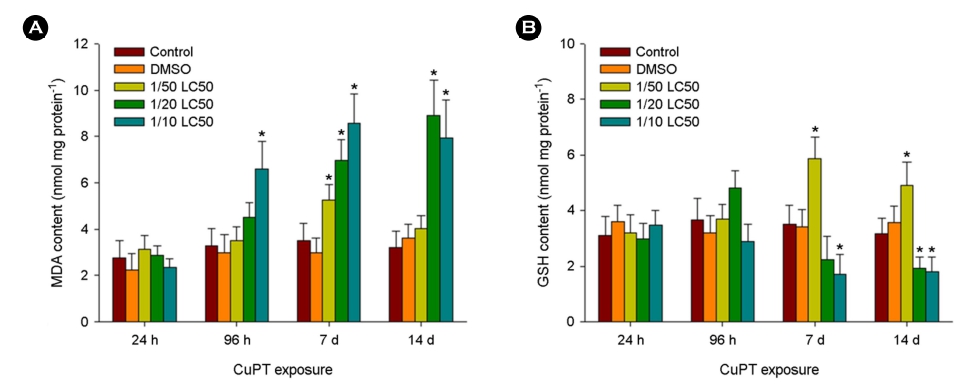
In marine polychaetes exposed to CuPT, significant increases in MDA content were observed in response to 1/10 LC50 value at 96 h, 1/50, 1/20, and 1/10 LC50 values at day 7, and 1/20 and 1/10 LC50 values at days 14, compared with the control (p < 0.05) (Fig. 2A).
Significantly elevated GSH contents were detected in response to 1/50 LC50 value on day 7 and 14, whereas significantly decreased GSH contents were observed after exposure to 1/10 LC50 value on day 7 and 1/20 and 1/10 LC50 values on day 14 (p < 0.05) (Fig. 2B).
3. Effects on the antioxidant defense system
CAT activity was significantly increased in the 1/50 LC50 CuPT-exposed polychaetes at 96 h and day 14, whereas significantly decreased activities were observed in response to 1/10 LC50 value on days 7 and 14 (p < 0.05) (Fig. 3A). SOD activity was significantly increased in response to 1/50 LC50 value at 24 h, 96 h, and day 14 and 1/20 LC50 value at 96 h, while significantly decreased activities were measured after exposure to 1/10 LC50 value at day 7 and 1/20 and 1/10 LC50 values at day 14 (p < 0.05) (Fig. 3B). GR activity was significantly increased by 1/50 and 1/10 LC50 values at 24 h and 1/50 and 1/20 LC50 values at 96 h, respectively, whereas its activities were significantly decreased by 1/20 and 1/10 LC50 values at day 7 and 1/50, 1/20, and 1/10 LC50 at day 14, respectively (p < 0.05) (Fig. 3C). The enzymatic activity of GPx was significantly increased by 1/20 LC50 value at 96 h, whereas significantly decreased activity was observed in response to 1/10 LC50 value on day 7 and 14 (p < 0.05) (Fig. 3D).

4. Effects on the GST activity
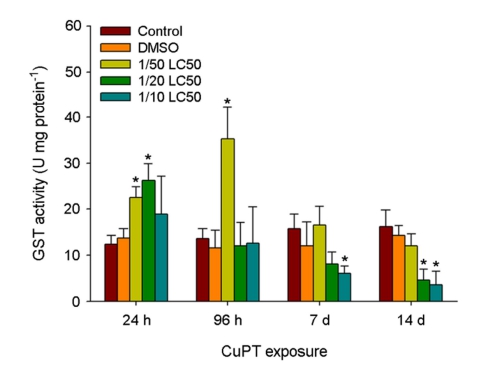
The enzymatic activities of GST were significantly elevated by 1/50 and 1/20 LC50 values at 24 h and 1/50 LC50 value at 96 h, whereas significantly decreased GST activities were observed in response to 1/10 LC50 value at day 7 and 1/20 and 1/10 LC50 values at day 14 (p < 0.05) (Fig. 4).
Discussion
Polychaetes are crucial ecosystem engineers because of their bioturbation activities, such as burrowing, ingestion, and defecation of sediment (Dean, 2008). In this study, the delay in the burrowing activity of P. aibuhitensis exposed to CuPT indicates a potential toxic effect of CuPT on basic physiology. Damage on muscle morphology, excessive oxidative stress, and fluctuations in neural activity, metabolism, and energy production upon CuPT would be the source of the negative effect on the burrowing behavior of polychaetes (Dorgan et al., 2007, 2008). Previously, similar dose-dependent delay in burrowing activity was observed in response to antifoulant ZnPT in P. aibuhitensis (Haque et al., 2020). A significant retardation in burrowing time was observed in the polychaete Hediste diversicolor exposed to 1,000 ng l-1 of diaze- pam for 28 days (Nogueira and Nunes, 2021).
The inhibition of AChE can decrease the hydrolysis of acetyl- choline (ACh) into choline and subsequently induces ACh accumulation at synapses (Nunes, 2011). In polychaetes, GABAergic neurotransmission plays a crucial role in behavior and locomotion (Biscocho et al., 2018). Thus, the AChE inhibition observed in this study demonstrates the interference of regular nerve communi- cation and induction of neurotoxicity by CuPT, as reflected by the delayed burrowing activity. In the mummichog Fundulus heteroclitus, exposure to 2 and 4 μg l-1 of CuPT significantly in- hibited AChE activity for 96 h (Mochida et al., 2008). Pyrithione-based antifoluant, ZnPT also inhibited AChE activity in the poly- chaete H. diversicolor exposed to 40 μg l-1 for 96 h (Nunes and Costa, 2019). To understand whether fluctuations in oxidative status and antioxidant defense system can be additional modulators on cholinergic damage, measurements were expanded to the responses of key regulators during oxidative stress in P. aibuhitensis.
The significant increase in intracellular MDA indicates CuPT-triggered severe lipid peroxidation in P. aibuhitensis. Lipid per- oxidation is a well-known consequence of oxidative stress. In detail, lipid peroxidation can be induced by reactive oxidants (e.g., hydroxyl radical, peroxides, superoxide, and singlet oxygen) through damage to the carbon-carbon double bonded polyunsaturated fatty acids of membrane lipids (Livingstone, 2001; Lesser, 2006). Previously, significant increase in intracellular MDA level was observed in zebrafish Danio rerio exposed to 32 and 64 μg l-1 of CuPT for 24 h (Almond and Trombetta, 2016). Similar increases in MDA content have been observed in aquatic animals exposed to other biofoulants. For example, exposure to 10 and 50 μg l-1 of ZnPT for 30 days significantly elevated intracellular MDA level in the olive flounder Paralichthys olivaceus (Min et al., 2019). A significant increase in MDA content in response to Sea-Nine was reported in this species (Eom et al., 2019).
Elevated GSH content is typically associated with increased defense against oxidative stress and its depletion is attributed to GSH consumption or direct GSH oxidation fostered by reactive oxidants (Winston and Di Giulio, 1991; Lushchak, 2011). Increased GSH level observed in the CuPT-exposed P. aibuhitensis would be explained as an adaptive response (neutralizing or detoxifying the reactive oxidants and peroxides) against oxidative stress. However, decreased GSH content can be suggested as drastic GSH con- sumption or inhibition of intracellular GSH synthesis through enhanced production of reactive oxidants by CuPT. This result suggests a mutual correlation between significantly increased lipid peroxidation and GSH synthesis or its depletion as a response against CuPT-triggered oxidative stress.
One of the main reactive oxidants, produced in cells in response to xenobiotics, is superoxide ion (O2•-), which can be catalyzed by the SOD enzyme into O2 and H2O2 (Lesser, 2006). To reduce the toxic H2O2, CAT enzyme prevents its accumulation by converting the H2O2 into H2O and O2 (Lushchak, 2011). Thus, both SOD and CAT enzymes play a robust antioxidant role by eliminating toxic oxidative elements from cells (Winston and Di Giulio, 1991; Lushchak, 2011). In this study, the significant increase in SOD and CAT activities in response to relatively low concentrations of CuPT suggests an activity for the rapid elimination of the reactive oxidants, whereas the significantly decreased activities of SOD and CAT enzymes by relatively high concentrations of CuPT indicates the accumulation of reactive oxidants, failure of reactive oxidants removal, and induction of severe oxidative stress (Lesser, 2006; Lushchak, 2011).
The antioxidant components, GR and GPx enzymes play a crucial role in reducing reactive oxidants, maintaining GSH levels, and minimizing oxidative stress in cells (Winston and Di Giulio, 1991; Lesser, 2006). GR enzyme is responsible for the reduction of glutathione disulfide (GSSG) to GSH, and GPx enzyme utilizes the GSH to degrade H2O2 into H2O (Lesser, 2006). GPx enzyme requires GSH as a co-factor to produce GSSG as a product (Livingstone, 2001). Significantly increased enzymatic activities of GR and GPx observed in relatively low concentrations of CuPT-exposed P. aibuhitensis indicate that both enzymes are strongly involved in the removal of reactive oxidants by the conjugation with GSH for elimination of oxidative toxicity. However, the significant decreases in the enzymatic activities of GPx and GR in response to relatively high concentrations of CuPT would be attributed to the accumu- lation of oxyradicals or H2O2 and failure of their removal as well as decreased efficacy of their conjugation process with GSH.
Phase II detoxification process is managed by several super- families of conjugation enzymes, which increase the solubility and reduce the toxicity of xenobiotics products transformed by phase I detoxification process. The phase II detoxification enzyme, GST uses GSH to detoxify xenobiotics to less toxic compounds (Sies, 1997). In P. aibuhitensis, the significant increase in GST activity would be regarded as an induction of cellular detoxification metab- olism to eliminate reactive oxidants through conjugation with GSH. In the case of decreased GST activity in response to relatively high concentrations of CuPT can be attributed to the substantial depletion of the substrate GSH and potential effects of CuPT on the cellular detoxification mechanism by overproduction of reactive oxidants (Sheehan et al., 2001; Lesser, 2006).
Conclusions
This is the first report exposing a marine polychaete to sublethal concentrations of the antifoulant CuPT. The overall results analyzed in this study show that CuPT could exert harmful effects on the cholinergic system, antioxidant defense system, and detoxification metabolism in marine polychaetes. The severity of the impairment of cholinergic parameters and antioxidant response was clearly associated with the concentration of CuPT. Although alternatives to TBT including CuPT has been introduced in antifouling paints due to the harmful effects of TBT on non-target marine species, results from this study suggest potential toxicity to the non-target animals close to that of the TBT-based antifoulants. In addition, the biomarker parameters employed in this study can be widely used as promising indicator of CuPT contamination in marine environments. However, more studies are needed to understand the mode of toxicity of CuPT in the other benthic communities at the ecological level.
- References
-
1. Almond KM, Trombetta LD. 2016. The effects of copper pyrithione, an antifouling agent, on developing zebrafish embryos. Ecotoxicology 25: 389-398.
-
2. Biscocho D, Cook JG, Long J, Shah N, Leise EM. 2018. GABA is an inhibitory neurotransmitter in the neural circuit regulating metamorphosis in a marine snail. Dev Neurobiol 78: 736-753.
-
3. Bonnard M, Romeo M, Amiard-Triquet C. 2009. Effects of copper on the burrowing behavior of estuarine and coastal inver- tebrates, the polychaete Nereis diversicolor and the bivalve Scrobicularia plana. Hum Ecol Risk Assess 15: 11-26.
-
4. Bradford MM. 1976. A rapid and sensitive method for the quanti- tation of microgram quantities of protein utilizing the prin- ciple of protein-dye binding. Anal Biochem 72: 248-254.
-
5. Dean HK. 2008. The use of polychaetes (Annelida) as indicator species of marine pollution: a review. Rev Biol Trop 56: 11-38.
-
6. Doose CA, Ranke J, Stock F, Bottin-Weber U, Jastorff B. 2004. Structure-activity relationships of pyrithiones-IPC-81 toxicity tests with the antifouling biocide zinc pyrithione and structural analogs. Green Chem 6: 259-266.
-
7. Dorgan KM, Arwade SR, Jumars PA. 2007. Burrowing in marine muds by crack propagation: kinematics and forces. J Exp Biol 210: 4198-4212.
-
8. Dorgan KM, Arwade SR, Jumars PA. 2008. Worms as wedges: effects of sediment mechanics on burrowing behavior. J Mar Res 66: 219-254.
-
9. Ellman GL, Courtney KD, Andres Jr. V, Feather-Stone RM. 1961. A new and rapid colorimetric determination of acetylcholin- esterase activity. Biochem Pharmacol 7: 88-95.
-
10. Eom H-J, Haque MN, Nam S-E, Lee D-H, Rhee J-S. 2019. Effects of sublethal concentrations of the antifouling biocide Sea-Nine on biochemical parameters of the marine polychaete Perinereis aibuhitensis. Comp Biochem Physiol C Toxicol Pharmacol 222: 125-134.
-
11. Fonseca TG, Auguste M, Ribeiro F, Cardoso C, Mestre NC, Abessa DMS, Bebianno MJ. 2018. Environmental relevant levels of the cytotoxic drug cyclophosphamide produce harmful effects in the polychaete Nereis diversicolor. Sci Total Environ 636: 798-809.
-
12. Goka K. 1999. Embryotoxicity of zinc pyrithione, an antidandruff chemical, in fish. Environ Res 81: 81-83.
-
13. Grunnet KS, Dahllof I. 2005. Environmental fate of the antifouling compound zinc pyrithione in seawater. Environ Toxicol Chem 24: 3001-3006.
-
14. Guthery E, Seal LA, Anderson EL. 2005. Zinc pyrithione in alcohol-based products for skin antisepsis: persistence of antimicrobial effects. Am J Infect Control 33: 15-22.
-
15. Haque MN, Nam S-E, Eom H-J, Kim S-K, Rhee J-S. 2020. Exposure to sublethal concentrations of zinc pyrithione inhibits growth and survival of marine polychaete through induction of oxi- dative stress and DNA damage. Mar Pollut Bull 156: 111276.
-
16. Kobayashi N, Okamura H. 2002. Effects of new antifouling com- pounds on the development of sea urchin. Mar Pollut Bull 44: 748-751.
-
17. Lesser MP. 2006. Oxidative stress in marine environments: bio- chemistry and physiological ecology. Annu Rev Physiol 68: 253-278.
-
18. Livingstone DR. 2001. Contaminated-stimulated reactive oxygen species production and oxidative damage in aquatic organ- isms. Mar Pollut Bull 42: 656-666.
-
19. Lushchak VI. 2011. Environmentally induced oxidative stress in aquatic animals. Aquat Toxicol 101: 13-30.
-
20. Min B-H, Saravanan M, Nam S-E, Eom H-J, Rhee J-S. 2019. Waterborne zinc pyrithione modulates immunity, biochemical, and antioxidant parameters in the blood of olive flounder. Fish Shellfish Immunol 92: 469-479.
-
21. Mochida K, Ito K, Harino H, Onduka T, Kakuno A, Fujii K. 2008. Early life-stage toxicity test for copper pyrithione and in- duction of skeletal anomaly in a teleost, the mummichog (Fundulus heteroclitus). Environ Toxicol Chem 27: 367-374.
-
22. Nogueira AF, Nunes B. 2021. Acute and chronic effects of diazepam on the polychaete Hediste diversicolor: Antioxidant, metabolic, pharmacologic, neurotoxic and behavioural mechanistic traits. Environ Toxicol Pharmacol 82: 103538.
-
23. Nunes B, Costa M. 2019. Study of the effects of zinc pyrithione in biochemical parameters of the Polychaeta Hediste diversicolor: evidences of neurotoxicity at ecologically relevant concen- trations. Environ Sci Pollut Res 26: 13551-13559.
-
24. Nunes B. 2011. The use of cholinesterases in ecotoxicology. Rev Environ Contam Toxicol 212: 29-59.
-
25. Reish DJ, Gerlinger TV. 1997. A review of the toxicological studies with polychaetous annelids. Bull Mar Sci 60: 584-607.
-
26. Rhee J-S, Lee Y-M, Hwang D-S, Won E-J, Raisuddin S, Shin K-H, Lee J-S. 2007. Molecular cloning, expression, biochemical characteristics, and biomarker potential of theta class glu- tathione S-transferase (GST-T) from the polychaete Neanthes succinea. Aquat Toxicol 83: 104-115.
-
27. Rhee J-S, Won E-J, Kim R-O, Choi B-S, Choi I-Y, Park GS, Shin K-H, Lee Y-M, Lee J-S. 2012. The polychaete, Perinereis nuntia ESTs and its use to uncover potential biomarker genes for molecular ecotoxicological studies. Environ Res 112: 48-57.
-
28. Schiff K, Brown J, Diehl D, Greenstein D. 2007. Extent and magni- tude of copper contamination in marinas of the San Diego region, California, USA. Mar Pollut Bull 54: 322-328.
-
29. Sheehan D, Foley DM, Dowd CA. 2001. Structure, function and evolution of glutathione transferases: implications for classi- fication of non-mammalian members of an ancient enzyme superfamily. Biochem J 360: 1-16.
-
-
31. Thomas KV. 1999. Determination of the antifouling agent zinc pyrithione in water samples by copper chelate formation and high performance liquid chromatography-atmospheric pres- sure chemical ionization mass spectrometry. J Chromatogr A 833: 105-109.
-
32. Winston GW, Di Giulio RT. 1991. Prooxidant and antioxidant mech- anisms in aquatic organisms. Aquat Toxicol. 19: 137-161.
-
33. Yebra DM, Kiil S, Johansen KD. 2004. Antifouling technology - past, present and future steps towards efficient and environmen- tally friendly antifouling coatings. Prog Org Coat 50: 75-104.
Copyright (C) 2024. The Korean Society of Marine Life Science