










JMLS 2022 December;7(2):94-101. 10.23005/ksmls.2022.7.2.94 Epub 2022 December 14
Copyright © 2022 by The Korean Society of Marine Life Science
Effects of Cochlodinium polykrikoides on Oxidative Status and Immune Parameters in the Marine Medaka Oryzias javanicus
Seong Duk Do; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
Yun Kyung Shin; Aquaculture Industry Research Division, South Sea Fisheries Research Institute, National Institute of Fisheries Science, Jeonnam 59780, Korea
Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea, Yellow Sea Research Institute, Incheon 22012, Korea
- Abstract
In this study, the marine medaka Oryzias javanicus was exposed to two concentrations of non-toxin-producing red tide dinoflagellate C. polykrikoides (1,000 and 2,000 cells ml-1) for 96 h, and the time-course biochemical responses of antioxidant and immunity parameters were analyzed in the liver tissue. Significant ichthyotoxicity with increasing cell concentrations of C. polykrikoides and exposure period was observed for 96 h. Opercular respiratory rate was lowered in marine medaka exposed to 2,000 cells ml-1 of C. polykrikoides. Intracellular malondialdehyde (MDA) content significantly elevated in response to both cell concentrations. In the case of glutathione (GSH) content, the levels were significantly elevated by 1,000 cells ml-1 of C. polykrikoides, but the contents significantly depleted upon exposure to 2,000 cells ml-1 of C. polykrikoides. Similarly, enzymatic activities of catalase (CAT) and superoxide dismutase (SOD) were increased by 1,000 cells ml-1 of C. polykrikoides, whereas their activities were lowered by 2,000 cells ml-1 of C. polykrikoides. Analysis of the two immunity parameters, alternative complement pathway and lysozyme, showed significantly lowered activities in 2,000 cells ml-1 of C. polykrikoides-exposed liver tissue. These biochemical effects of C. polykrikoides on marine medaka would be helpful for understanding its acute effects in marine fish.
Keywords: Cochlodinium polykrikoides; Red tide Marine medaka Oxidative stress Immunity
Correspondence to: Jae-Sung Rhee; Department of Marine Science, College of Natural Sciences, Incheon National University, Incheon 22012, Korea
- Received
- 20 August 2022;
- Revised
- 7 September 2022;
- Accepted
- 22 November 2022.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

Introduction
Harmful algal blooms (HABs) are known as to be occurred through excessive growth of several microscopic algae or algae-like bacteria in aquatic ecosystems, although their blooms are natural phenomena (Landsberg, 2002). HABs are strongly asso- ciated with direct damage and/or their toxins to aquaculture and fisheries and are responsible for mass mortality of fish and shellfish (Hallegraeff, 2003). Detrimental effects of HABs on the econom- ically important aquaculture and fisheries have been highlighted worldwide (Whyte et al., 2001; Kudela and Gobler, 2012). Dinofla- gellates are abundant marine planktons with high diversity and are also crucial food sources in aquatic tropic levels (Shumway, 1990). HABs induced by dinoflagellates are often called red tides in marine ecosystem. In particular, several species of dinoflagellates can produce ichthyotoxic compounds even at low cell densities and induce numerous harmful effects through bioaccumulation in different trophic levels including marine mammals and human (Landsberg, 2002; Hallegraeff, 2003; Mindy et al., 2010). Several physical and physiological damages were reported in fish species exposed to dinoflagellate. For example, direct physical contact of C. polykrikoides with gill tissues and epithelial hyperplasia was observed in fish (Gobler et al., 2008). Detrimental effects such as gill clogging, anoxia, convulsions, swelling and necrosis of the lamellar epithelium, osmoregulatory disruption, and/or excess mucus secretion have been observed in numerous fish species exposed to dinoflagellates (Kim et al., 1999; Chen and Chou, 2001; Cembella et al., 2002; Landsberg, 2002; Band-Schmidt et al., 2003; Shin et al., 2019).
The unarmored, chain-forming mixotrophic dinoflagellate Cochlodinium polykrikoides is one of the most HAB-forming species with a wide range of distribution in the world (Whyte et al., 2001; Gárate-Lizárraga et al., 2004; Anton et al., 2008; Curtiss et al., 2008; Kudela et al., 2008; Tomas and Smayda, 2008). The genus Cochlodinium is known to be well adapted to environmental challenges, such as daily or seasonal fluctuations in temperature, salinity, irradiation, and nutrient (Kim et al., 2004; Gobler et al., 2008; Kudela et al., 2008; Mulholland et al., 2009). Apart from the toxins produced from Cochlodinium species, the mechanism of ichthyotoxicity of C. polykrikoides has been consistently sub- stantiated. C. polykrikoides can cause gill injury, hyperventilation, and respiratory impairment through direct attachment on the gill tissues of fish (Shin et al., 2019). Several non-toxin-producing Cochlodinium species are known to generate reactive oxygen species (ROS) such as superoxide anions and hydrogen peroxide that would be responsible for direct or indirect ichthyotoxicity (Kim et al., 1999; Marshall et al., 2002; Kim et al., 2004; Liu et al., 2007). In addition, a cocktail of highly reactive ROS-like compounds, mucus-like substances or biologically active metabolites released from Cochlodinium species were suggested as additives to ichthyo- toxicity (Kim et al., 2002; Tang and Gobler, 2009).
In general, fish undergo immunological fluctuations in response to environmental stressors. However, studies on HAB-triggered immunity response are scarce. Several immunity parameters such as measurement of enzymatic activity of complement and lyso- zyme have been used to reflect the health status and homeostasis of fish. The complement system has a crucial role in innate immunity with a wide range of immune recognition capabilities. Measurement of lysozyme activity has been consistently used for monitoring the potential impact of exogenous stress on innate immunity. To minimize the detrimental effects of Cochlodinium blooms and subsequent economic losses, it is crucial to under- stand the precise ichthyotoxicity mechanism, but there is limited knowledge on the actual effects of C. polykrikoides on fish liver, particularly the modulations of the antioxidant defense system and immunity.
In this study, we analyzed acute effects of C. polykrikoides on the liver tissue of the marine model fish, Oryzias javanicus. Although potential effects of C. polykrikoides have been reported in marine fish (e.g., red seabream), there is no information on the effects in small marine model fish. To estimate potential detri- mental effects, biochemical assays for antioxidant defense system and innate immunity were performed with 1,000 and 2,000 cells ml-1 of C. polykrikoides in this study. Intracellular contents of malondialdehyde (MDA) and glutathione (GSH) were monitored to confirm evidence for C. polykrikoides-mediated modulation in re- dox homeostasis. In addition, GSH-mediated antioxidant responses, as well as two immunity parameters, alternative complement pathway (ACH50) and lysozyme, were analyzed in the liver tissue after exposure to 1,000 and 2,000 cells ml-1 of C. polykrikoides. Our findings can help to understand C. polykrikoides toxicity in marine fish by providing biochemical responses for the threats of HABs.
Material and Methods
1. Experimental animals
Animal handling and experimental procedures were approved by the Animal Welfare Ethical Committee and the Animal Ex- perimental Ethics Committee of the Incheon National University (Incheon, South Korea). The marine medaka Oryzias javanicus used in this study were maintained in artificial seawater (TetraMarine Salt Pro, Cincinnati, OH, USA; 30 ± 0.5 practical salinity unit; 6.01± 0.76 mg O2 l-1) at 20 ± 1℃ under a 14/10 h L/D cycle. They were fed frozen mosquito larvae and an artificial diet twice a day until satiation.
A laboratory culture of C. polykrikoides was established in our previous study (Shin et al., 2019). It was grown at 20℃ in GSe medium dissolved with filtered seawater with a 14/10 h L/D cycle under a 120 μmol photons m-2 s-1 light source. Subculture was performed with an inoculation of approximately 50~60 cells ml-1 into 10 ℓ GSe medium before experiment. Cell number of C. polykrikoides was counted in a Leica DMLA microscope (Leica Microsystems, Wetzlar, Germany) equipped with UV epifluore- scence and an AxioCam HRc camera (Zeiss, Göttingen, Germany) using a Sedgwick-Rafter counting chamber (VWR, Langenfeld, Germany). For exposure, C. polykrikoides was harvested between the exponential and early stationary phases to adjust final con- centrations equivalent to approximately 10,000 C. polykrikoides cells ml-1 before treatment.
2. Analysis of acute effects of C. polykrikoides
During the survival rate measurement, fish were not pooled and one fish was reared on one experimental tank. Additional three-day acclimation was conducted in the experimental tanks to diminish potential stress on marine medaka. To measure sublethal ranges, 30 marine medaka were individually exposed to control (GSe medium) 1,000, 2,000, 3,000, 4,000, and 5,000 cells ml-1 of C. polykrikoides for 96 h. The fish were not fed during exposure. No marine medaka died for 96 h in the control group. Survival rates were monitored at 0, 3, 6, 12, 24, 48, and 96 h in the control and exposed marine medaka. The opercular respiratory rate of 1,000 and 2,000 cells ml-1-exposed marine medaka was monitored in a transparent chamber. Ten marine medaka in each concen- tration were moved into the chamber and the breath beats for each marine medaka were recorded for 5 min.
3. C. polykrikoides exposure and sample pre- paration
Sixty-three marine medaka per each concentration were dis- tributed into three groups and exposed to each concentration of C. polykrikoides. Three marine medaka per each concentration were collected at 0, 3, 6, 12, 24, 48, and 96 h. To sample liver tissue, individuals were anesthetized by immersion in a solution of tricaine methanesulfonate (Sigma-Aldrich, Inc. St. Louis, MO, USA). Each marine medaka was dissected to obtain liver tissues. Three liver samples from each subgroup were pooled for analysis, and the three pooled specimens were analyzed in triplicate.
4. Biochemical analysis
Pooled liver tissues were homogenized in a cold buffer (20 mM Tris, 150 mM NaCl, 10 mM β-mercaptoethanol, 20 μM leupeptin, 2 μM aprotinin, and 100 μM benzamidine) and centrifuged for 30 min at 30,000 × g at 4℃. After heat-denaturation of the super- natants for 15 min at 75℃, thiobarbituric acid reactives (TBARs) were measured at 535 nm with a Thermo Varioskan Flash spectro- photometer (Thermo Fisher Scientific, Tewksbury, MA, USA) using malonaldehyde bis (MDA; tetrametoxypropan, Sigma-Aldrich, Inc.) as standard. The total content of lipid peroxidation compounds was calculated as nM of MDA per gram of liver tissue. Total soluble protein was determined with the Bradford method.
Intracellular GSH content in the pooled liver tissues was analyzed using a Glutathione Assay Kit (Catalog No. CS0260; Sigma-Aldrich, Inc.). The samples were washed with 0.9% NaCl and were hom- ogenized in trichloroacetic acid with a Teflon homogenizer at a ratio of 1:20 (w/v). The homogenate was centrifuged at 3,000 × g for 10 min at 4℃ and supernatant was collected for the GSH assay according to the manufacturer's protocol. The GSH content was measured at an absorbance of 420 nm with a spectro- photometer, and the standard curves were generated with GSH equivalents (0, 150, and 350 μM).
Enzymatic activities of CAT and SOD were analyzed with enzy- matic methods using SOD Assay Kit (Catalog No. 19160; Sigma-Aldrich Chemie, Switzerland) and Catalase Assay Kit (Catalog No. CAT100; Sigma-Aldrich, Inc.), respectively. The CAT and SOD activities were calculated at an absorbance of 440 nm or 520 nm at 25℃ with a spectrophotometer, respectively.
The ACH50 activity of C. polykrikoides-exposed marine medaka was analyzed using sheep red blood cells (SRBCs; 1.5×106 cells, National Institute of Toxicological Research, South Korea) following a previous method (Nam et al., 2020). Ethylene glycol tetraacetic acid (EGTA)-Mg-gelatin-veronal buffer (GVB; pH 7.2), was prepared before the assay by mixing gelatin (0.1%), EGTA (10 mmol · l-1), and MgCl2 (10 mmol · l-1) and 25 μL of 6% serum was incubated in a 96-well plate with 5 μl of SRBCs and 10 mM phenol red-free Hank's buffer. The plate was incubated for 90 min at 25°C and 1 ml of 10 mM EDTA was added to the solution to stop the hemolytic reaction. The optical density of the mixture was scanned at a wavelength of 414 nm on the spectrophotometer. The volume of serum complement triggering 50% lysis of SRBCs (ACH50; U · ml-1) was measured.
The lysozyme activity of C. polykrikoides-exposed marine medaka was measured using a turbidimetric assay in a 96-well plate (Ellis, 1990). A substrate prepared with 0.3 mg · ml-1 lysozyme-sensitive bacterium Micrococcus lysodeikticus (ATCC No. 4698, Sigma-Aldrich) and 0.05 M sodium phosphate buffer (pH 6.2) was employed for analyzing lysozyme activity. Samples were examined in triplicate for statistical assurance. For each test, 10 μl of marine medaka sample and 200 μl of M. lysodeikticus suspension were mixed and incubated at room temperature. After 5 min of in- cubation, the absorbance of the mixture was read at a wavelength of 450 nm on the spectrophotometer.
5. Statistical analyses
All data were analyzed using the statistical software package SPSS (ver. 17.0, SPSS Inc., Chicago, IL, USA) and expressed as mean ± standard deviation (S.D.). A two-way analysis of variance (ANOVA) was employed to investigate the effect of C. polykrikoides on the results of antioxidant defense system and immunity. If significant differences were calculated by the ANOVA test, an additional Tukey test was used to confirm whether C. polykrikoides treatments and time periods were significantly different. Results with p < 0.05 were considered significant.
Results
1. Survival and respiration rate
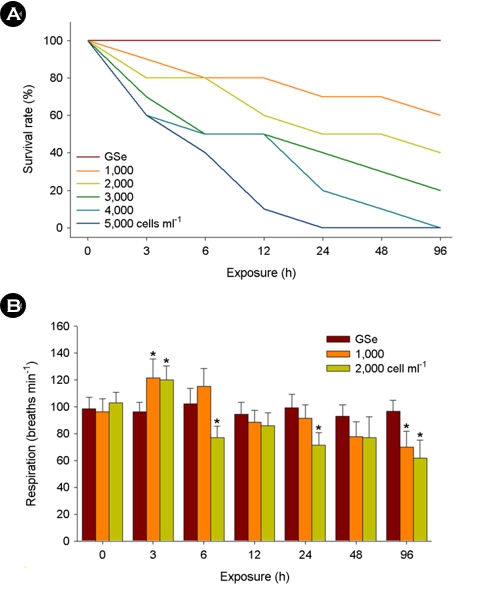
For exposure of 1,000 and 2,000 cells ml-1 of C. polykrikoides, no marine medaka died for 96 h in response to GSe medium, while concentration-dependent mortality was observed upon 1,000~5,000 cells ml-1 of C. polykrikoides (Fig. 1A).
The opercular respiratory rates or marine medaka were signifi- cantly elevated at 3 h in both 1,000 and 2,000 cells ml-1 of C. polykrikoides (p < 0.05), but the rates were significantly lowered at 6, 24, and 96 h in 2,000 cells ml-1 of C. polykrikoides-exposed marine medaka (p < 0.05) (Fig. 1B). No significant modulation of the opercular respiratory rate was detected in response to GSe medium (p > 0.05).
2. Malondialdehyde and glutathione content
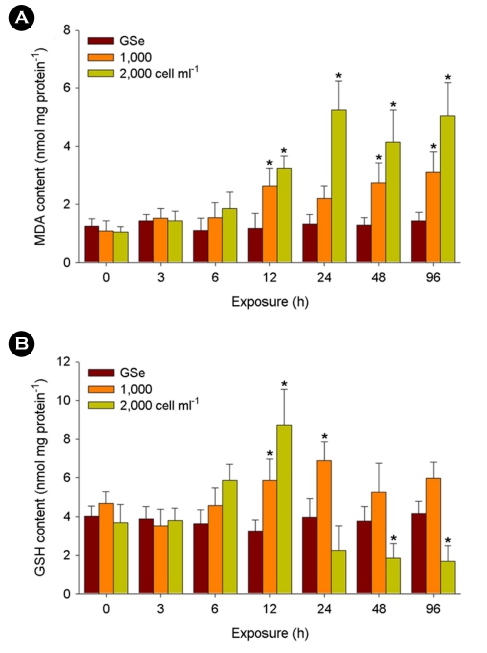
Significant increases in MDA content were detected in the liver tissues exposed to 1,000 cells ml-1 of C. polykrikoides at 12, 48, and 96 h and 2,000 cells ml-1 at 12, 24, 48, and 96 h (p < 0.05) (Fig. 2A). No significant change was analyzed in response to GSe medium (p > 0.05).
In the case of GSH content, 1,000 cells ml-1 of C. polykrikoides exposure resulted in its significantly higher levels at 12 and 24 h (p < 0.05), while the content significantly depleted after 48 h of exposure to 2,000 cells ml-1 (p < 0.05) (Fig. 2B). No significant change was analyzed in response to GSe medium (p > 0.05).
3. Antioxidant defense parameter
In the liver tissue exposed to 1,000 cells ml-1 of C. polykrikoides, CAT activity significantly elevated at 24, 48, and 96 h (p < 0.05), while the activity was significantly lowered by 2,000 cells ml-1 at 48 and 96 h after significantly higher levels measured at 12 and 24 h (p < 0.05) (Fig. 3A). No significant change was analyzed in response to GSe medium (p > 0.05).
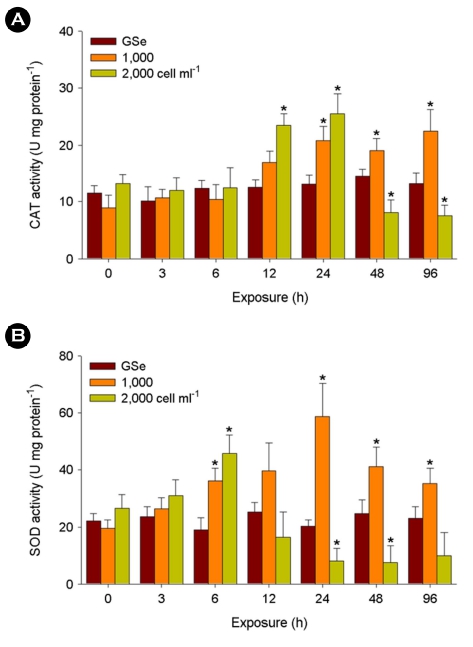
When exposed to 1,000 cells ml-1 of C. polykrikoides, signifi- cantly higher activity of the SOD enzyme was detected at 6, 24, 48, and 96 h (p < 0.05) (Fig. 4A). In addition, a significantly higher level of SOD activity was measured in the 2,000 cells ml-1 of C. polykrikoides-exposed liver tissue at 6 h (p < 0.05). The activity of SOD enzyme significantly lowered with exposure to 2,000 cells ml-1 of C. polykrikoides at 24 and 48 h (p < 0.05). No significant change was analyzed in response to GSe medium (p > 0.05).
4. Immune parameter
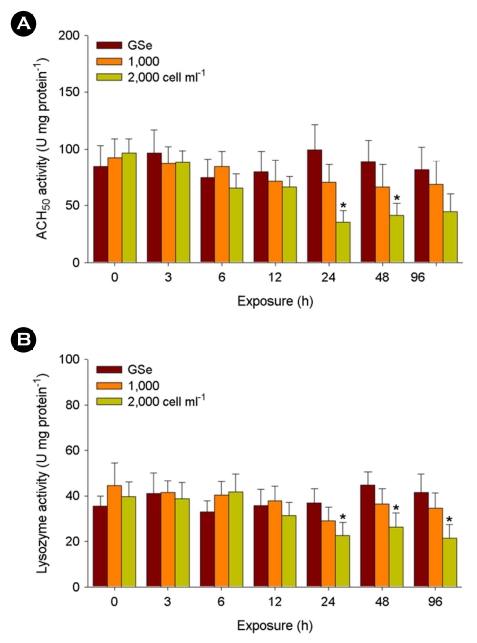
Significant decreases in ACH50 activity were observed in the liver tissues exposed to 2,000 cells ml-1 of C. polykrikoides at 24 and 48 h (p < 0.05) (Fig. 4A). No significant change was analyzed in response to GSe medium and 1,000 cells ml-1 of C. polykrikoides (p > 0.05).
Similarly, 2,000 cells ml-1 of C. polykrikoides exposure resulted in its significantly lower levels at 24, 48, and 96 h (p < 0.05), while no significant change was analyzed in response to GSe medium and 1,000 cells ml-1 of C. polykrikoides (p > 0.05) (Fig. 4B).
Discussion
The result of survival rate revealed that the cell concentration and exposure period of C. polykrikoides was critical for the ichthyo- toxicity in marine medaka. Similar concentration ranges of C. polykrikoides responsible for mortality were reported in multiple fish species (Gobler et al., 2008; Shin et al., 2019; Haque et al., 2021). In marine medaka, C. polykrikoides significantly modulated their respiration rates in a short time course. Thus, mortality observed in marine medaka would be associated with alteration of the gill epithelium structure, disruption of ionic homeostasis, and suffocation through C. polykrikoides attachment. Thus, the increased mortality would be resulted from failure in gill function and respiration through direct ingestion and attachment/aggregation of C. polykrikoides, as we used a non-toxin-producing species. Detrimental effects on gill tissues of fish is critical for mortality, as gills are a crucial organ for absorption and transport of oxygen into whole body.
To date, negative physical and physiological damages induced by C. polykrikoides have been investigated in fish (Gobler et al., 2008), but limited information is available on molecular and biochemical effects of C. polykrikoides exposure. Treatment to C. polykrikoides significantly elevated MDA contents and the oxi- dative stress parameter, GSH content in marine medaka. Malon- dialdehyde is one of the byproducts of lipid peroxidation. Sub- sequently, the measurement of intracellular MDA content has been widely employed as a promising biomarker for assessing oxidative stress in fish (Lushchak, 2011). Therefore, increased MDA content observed in marine medaka indicated that treatment to C. polykrikoides may induce production of oxidative stressors (e.g., free radicals or pro-oxidant molecules) that can further trigger an induction of lipid peroxidation. In the flatfish Paralichthys olivaceus exposed to C. polykrikoides, concentration-dependent increase in lipid peroxidation was observed (Kim et al., 1999).
Glutathione is an important non-enzymatic antioxidant as a crucial protector against cellular redox-cycling fluctuations by exogenous oxidative stressors (Lushchak and Bagnyukova, 2006). The significant increase of GSH levels observed in marine medaka suggests a synthesis of new GSH that may be needed to maintain its required levels for the increase of the resistance capacity against oxidative stress as a free radical scavenger. However, prolonged depletion of GSH content can lead to vulnerability to oxidative stress and lethal damage (Townsend et al., 2003). The strongly modulated respiration rate would be considered a factor for intracellular oxidative stress, as low oxygen-triggered lipid peroxidation and subsequent oxidative stress has been well-characterized in fish (Lushchak and Bagnyukova, 2006).
The two antioxidant parameters, CAT and SOD enzymes are the first line of antioxidant defense upon oxidative stress. In detail, SOD converts two superoxide radicals (O2-•) into hydrogen peroxide (H2O2) and CAT decomposes H2O2 into water (H2O) and molecular oxygen (O2), resulting in the diminishment of the detrimental effects of free radicals (Lesser, 2006; Lushchak, 2011). Thus, in- crease enzymatic activities of CAT and SOD observed in marine medaka suggest stress response and adaptive metabolism against oxidative stress. In the gill cell line RTgill-W1 exposed to the raphidophyte Chattonella marina, significantly increased SOD enzyme activity was observed (Dorantes-Aranda et al., 2015). However, their parallel reduction of CAT and SOD enzymes may be explained as failure in regulating oxidative homeostasis.
The immune parameters, ACH50 and lysozyme play a crucial role in innate immunity, immunosuppression, and homeostasis in fish (Boshra and Sunyer, 2006; Saurabh and Sahoo, 2008). The significant decreases in enzymatic activities of ACH50 and lysozyme measured in marine medaka represent C. polykrikoides-triggered immune suppression. Since the liver is the important organ that synthesizes complement components (Boshra and Sunyer, 2006), the significantly decreased ACH50 activity implies a potential inter ruption in liver function. In fish, macrophages, neutrophils, and monocytes release lysozymes that directly lyse or degrade pathogen cell walls (Magnadóttir, 2006). Previously, the dinofla- gellate Alexandrium affine (6,000 and 7,000 cells ml-1) induced immunosuppression by significant decreases in lysozyme activity and total Ig level in the gill and liver tissues of red seabream, Pagrus major (Haque et al., 2021). Therefore, the decreased activity of lysozyme represents significant immune suppression and poten- tially higher vulnerability of the marine medaka to diseases and exogenous pathogens and may allow unlimited inflammation.
In summary, all the parameters of the antioxidant defense system and immunity analyzed in the liver tissue support oxidative stress induction and immunosuppression by C. polykrikoides in marine medaka. Our results also suggest that even environmental con- centrations of HABs can inhibit respiration rate and may induce mass mortality in marine fish. Detection of early biological signals of aquatic animals in response to HABs will be useful to predict the occurrence of HABs and establish prevention plans in aqua- culture and fisheries.
- References
-
1. Anton A, Teoh PL, Mohd-Shaleh SR, Mohammad-Noor N. 2008. First occurrence of Cochlodinium blooms in Sabah, Malaysia. Harmful Algae 7: 331-336.
-
2. Band-Schmidt CJ, Lechuga-Deveze CH, Kulis DM, Anderson DM. 2003. Culture studies of Alexandrium affine (Dinophyceae), a non-toxic cyst forming dinoflagellate from Bahía concepción, Gulf of California. Bot Mar 46: 44-54.
-
3. Boshra H, Li J, Sunyer JO. 2006. Recent advances on the com- plement system of teleost fish. Fish Shellfish Immunol 20: 239-262.
-
4. Cembella AD, Quilliam MA, Lewis NI, Bauder AG, Dell'Aversano C, Thomas K, Jellett J, Cusack RR. 2002. The toxigenic marine dinoflagellate Alexandrium tamarense as the probable cause of mortality of caged salmon in Nova Scotia. Harmful Algae 1: 313-325.
-
5. Chen CY, Chou HN. 2001. Ichthyotoxicity studies of milkfish Chanos chanos fingerlings exposed to a harmful dinoflagellate Alexandrium minutum. J Exp Mar Biol Ecol 262: 211-219.
-
6. Curtiss CC, Langlois GW, Busse LB, Mazzillo F, Silver MW. 2008. The emergence of Cochlodinium along the California Coast (USA). Harmful Algae 7: 337-346.
-
7. Dorantes-Aranda JJ, Seger A, Mardones JI, Nichols PD, Hallegraeff GM. 2015. Progress in understanding algal bloom-mediated fish kills: The role of superoxide radicals, phycotoxins and fatty acids. PLoS One 10: e0133549.
-
8. Ellis AE. 1990. Lysozyme assays. In: Stolen JS, Fletcher TC, Anderson DP, Roberson BS, Van Muiswinkel WB. (Eds.), Techniques in Fish Immunology. SOS Publications, Fair Haven NJ, pp 101-103.
-
9. Gárate-Lizárraga I, López-Cortes DJ, Bustillo-Guzmán JJ, Hemández-Sandoval F. 2004. Blooms of Cochlodinium polykrikoides (Gymnodiniaceae) in the Gulf of California, Mexico. Rev Biol Trop 52: 51-58.
-
10. Gobler CJ, Berry DL, Anderson OR, Burson A, Koch F, Rodgers BS, Moore LK, Goleski JA, Allam B, Bowser P, Tang Y, Nuzzi R. 2008. Characterization, dynamics, and ecological impacts of harmful Cochlodinium polykrikoides blooms on eastern Long Island, NY, USA. Harmful Algae 7: 293-307.
-
11. Hallegraeff GM. 2003. Harmful algal blooms: a global overview, in Hallegraeff GM, Anderson DM, Cembella AD. (Eds.), Manual on harmful marine microalgae. UNESCO, Paris, France, pp 25-49.
-
12. Haque MN, Nam S-E, Shin YK, Rhee J-S. 2021. The dinoflagellate Alexandrium affine acutely induces significant modulations on innate immunity, hepatic function, and antioxidant defense system in the gill and liver tissues of red seabream. Aquat Toxicol 240: 105985.
-
13. Kim CS, Lee SG, Kim HG, Jung J. 1999. Reactive oxygen species as causative agents in the ichthyotoxicity of the red tide dinoflagellate Cochlodinium polykrikoides. J Plankton Res 21: 2105-2115.
-
14. Kim D, Oda T, Muramatsu T, Kim D, Matsuyama Y, Honjo T. 2002. Possible factors responsible for the toxicity of Cochlodinium polykrikoides, a red tide phytoplankton. Comp Biochem Physiol C 132: 415-423.
-
15. Kim D-I, Matsuyama Y, Nagasoe S, Yamaguchi M, Yoon Y-H, Oshima Y, Imada N, Honjo T. 2004. Effects of temperature, salinity and irradiance on the growth of the harmful red tide dinoflagellate Cochlodinium polykrikoides Margalef (Dino- phyceae). J Plankton Res 26: 61-66.
-
16. Kudela RM, Ryan JP, Blakely MD, Lane JQ, Peterson TD. 2008. Linking the physiology and ecology of Cochlodinium to better understand harmful algal bloom events: a comparative approach. Harmful Algae 7: 278-292.
-
17. Kudela R, Gobler C. 2012. Harmful dinoflagellate blooms caused by Cochlodinium sp.: global expansion and ecological strategies facilitating bloom formation. Harmful Algae 14: 71-86.
-
18. Landsberg JH. 2002. The effects of harmful algal blooms on aquatic organisms. Rev Fish Sci 10: 113-390.
-
19. Lesser MP. 2006. Oxidative stress in marine environments: bio- chemistry and physiological ecology. Annu Rev Physiol 68: 253-278.
-
20. Liu W, Au DW, Anderson DM, Lam PK, Wu RS. 2007. Effects of nutrients, salinity, pH and light:dark cycle on the production of reactive oxygen species in the alga Chattonella marina. J Exp Mar Biol Ecol 346: 76-86.
-
21. Lushchak VI, Bagnyukova TV. 2006. Effects of different environ- mental oxygen levels on free radical processes in fish. Comp Biochem Physiol B 144: 283-289.
-
22. Lushchak VI. 2011. Environmentally induced oxidative stress in aquatic animals. Aquat Toxicol 101: 13-30.
-
-
24. Marshall J-A, Hovenden M, Oda T, Hallegraeff GM. 2002. Photo- synthesis does influence superoxide production in the ichthyotoxic alga Chattonella marina (Raphidophyceae). J Plankton Res 24: 1231-1236.
-
25. Mindy LR, Steve LM, Ebrahim AJ, Anbiah R, Donald MA. 2010. The catastrophic 2008-2009 red tide in the Arabian Gulfregion, with observations on the identification and phylogeny of the fish killing dinoflagellate Cochlodinium polykrikoides. Harmful Algae 9: 163-172.
-
26. Mulholland MR, Morse RE, Boneillo GE, Bernhardt PW, Filippino KC, Procise LA, Blanco-Garcia JL, Marshall HG, Egerton TA, Hunley WS, Moore KA, Berry DL, Gobler CJ. 2009. Under- standing causes and impacts of the dinoflagellate, Cochlo- dinium polykrikoides, blooms in the Chesapeake Bay. Estuar Coast 32: 734-747.
-
27. Nam S-E, Saravanan M, Rhee J-S. 2020. Benzo[a]pyrene constrains embryo development via oxidative stress induction and modulates the transcriptional responses of molecular bio- markers in the marine medaka Oryzias javanicus. J Environ Sci Health A 55: 1050-1058.
-
28. Saurabh S, Sahoo PK. 2008. Lysozyme: an important defence molecule of fish innate immune system. Aquac Res 39: 223-239.
-
29. Shin Y-K, Nam S-E, Kim WJ, Seo DY, Kim Y-J, Rhee J-S. 2019. Red tide dinoflagellate Cochlodinium polykrikoides induces sig- nificant oxidative stress and DNA damage in the gill tissue of the red seabream Pagrus major. Harmful Algae 86: 37-45.
-
30. Shumway SE. 1990. A review of the effects of HABs on shellfish and aquaculture. J World Aquacult Soc 21: 65-104.
-
31. Tang YZ, Gobler CJ. 2009. Characterization of the toxicity of Cochlodinium polykrikoides isolates from Northeast US estuaries to finfish and shellfish. Harmful Algae 8: 454-462.
-
32. Tomas CR, Smayda TJ. 2008. Red tide blooms of Cochlodinium polykrikoides in a coastal cove. Harmful Algae 7: 308-317.
-
33. Townsend DM, Tew KD, Tapiero H. 2003. The importance of glu- tathione in human disease. Biomed Pharmacother 57: 145-155.
-
34. Whyte JNC, Haigh N, Ginther NG, Keddy LJ. 2001. First record of blooms of Cochlodinium sp. (Gymnodiniales, Dinophyceae) causing mortality to aquacultured salmon on the west coast of Canada. Phycologia 40: 298-304.
Copyright (C) 2024. The Korean Society of Marine Life Science