










JMLS 2022 December;7(2):61-73. 10.23005/ksmls.2022.7.2.61 Epub 2022 December 14
Copyright © 2022 by The Korean Society of Marine Life Science
Application of Stable Isotope Analysis for Aquaculture Organisms
원은지; 한양대학교 해양대기과학연구소,한양대학교 해양융합과학과
윤희영; 한양대학교 해양대기과학연구소
최보형; 국립수산과학원 중앙내수면연구소
신경훈; 한양대학교 해양대기과학연구소,한양대학교 해양융합과학과
- Abstract
안정동위원소 분석 기법(Stable isotope analysis, SIA)은 환경과학, 생태학, 지구생물화학, 법의학, 고고학 등 다양한 연구 분야에 활용되고 있다. 본 총설에서는 수산 및 양식 연구에 안정동위원소 비 분석 기법을 활용하기 위해 필요한 배경 지식을 소개하고자 한다. 특히, 자연 값(natural abundance)을 이용하는 연구에 초점을 두었고 원소가 생물의 조직으로 통합되는 과정에서 발생하는 분별작용(동위원소 비의 변화)에 대한 원리와 안정동위원소 비가 유용한 도구로서 어떤 목적으로 생태, 환경학 분야에 이용되는지, 나아가 수산 및 양식 분야에 활용 가능한 예들을 제시하고자 한다. 본 논문을 통한 안정동위원소 분야의 이해로 향후 수산학 및 양식 연구에서 안정 동위원소 비의 다양한 활용이 기대된다.
Stable isotope analysis (SIA) is being used in various research fields including environmental science, ecology, biogeochemistry, forensics, and archeology. In this paper, for the purpose of enhancing applications and utilizations stable isotope analysis techniques to aquaculture research, we would like to introduce the background knowledge necessary to utilize stable isotope analysis techniques. In particular, with a focus on the approach using natural abundance, the principle of fractionation (change in isotope ratio) that occurs in the process of the integration of elements into biological tissues and how stable isotope ratios are determined by fractionation. This paper is intended to suggest whether SIA is used as a valuable tool in the fields of ecology and environmental science. With the understanding of the field of stable isotopes through this paper, various applications of stable isotope ratios are expected in fisheries science and aquaculture research in the future.
Keywords: Compound specific isotopes(화합물 안정동위원소) Food quality(먹이질) Isotope physiology(동위원소생리학) Isotope ecology(동위원소생태학) Geographical authentication(산지 판별)
Correspondence to: Eun-Ji Won; Institute of Ocean and Atmospheric Sciences (IOAS), Hanyang University, Ansan 15588, Korea
- Received
- 8 November 2022;
- Revised
- 10 November 2022;
- Accepted
- 22 November 2022.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
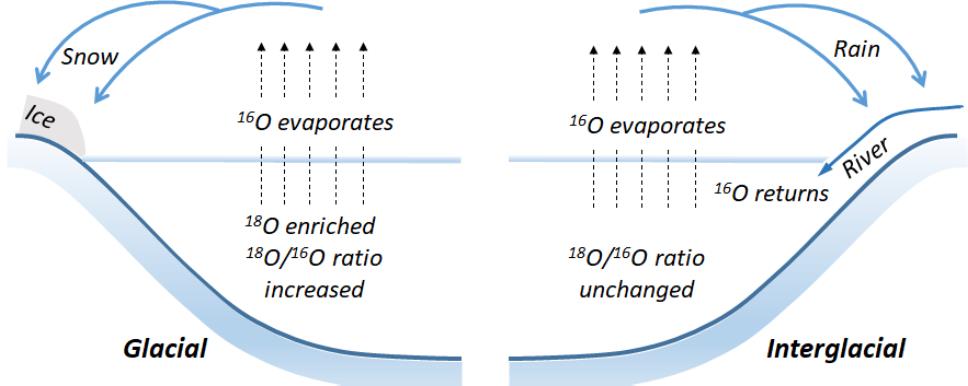
동위원소는 원자 번호가 같지만 구성하고 있는 질량수가 다른 원소로 양성자의 수가 같고 중성자수가 다른 핵 종을 뜻한다(Ben-David and Flaherty, 2012). 그 중 방사 붕괴를 하지 않는 동위 원소를 안정동위원소(stable isotope)라고 하는데 수소(3H, D), 탄소(13C), 질소(15N), 산소(18O), 황(34S) 등의 원소가 이에 해당한다. 동위원소 질량분석기(isotope ratio mass spectrometry)를 이용한 동위원소 비 분석은 동위원소를 이용할 수 있는 다양한 연구에 사용되어왔다. 안정동위원소는 방사성동위원소를 대체하며 의학, 생물학, 화학, 물리학 등의 분야에서 이용되어 왔으며, 그 예로, 동위원소를 추적자(e.g., 13C)로 이용함으로써 세포 내 물질의 생성, 반응과 속도, 또는 반응이 일어나는 조직을 추적하는 등 생물학적, 화학적 반응을 해석하는 연구에 유용하게 활용되고 있다(Gilbert et al., 2012; Min et al., 2019). 환경과학 및 생태학 연구에서는 자연에 존재하는 동위원소 비(natural abundance)를 이용한 연구도 활발하게 이루어지고 있다. 그 예로 산소의 경우, 자연 상태의 존재 비는 99.76% 이상이 질량수 16, 0.04% 정도는 질량수 17인 산소이며 0.2%가 질량수 18인 산소로 존재한다. 그러나 산소가 이용되는 물리 화학적인 반응에서 가벼운 16O가 반응에 필요한 에너지가 작아 반응이 빠르고 많이 일어나게 되고 이러한 질량 분별효과(fractionation)를 이용해 반응이 일어난 조건(즉, 환경)을 설명할 수 있다. 대표적인 반응으로 물의 증발에서는 상대적으로 무거운 동위원소인 18O가 수증기에 덜 포함되게 되는데, 극지의 빙하와 심해저의 유공충(foraminifera)에서 분석된 산소 동위원소의 자연 존재 비(16O와 18O의 비율)를 이용해 증발이 일어날 당시의 고해양(paleo-ocean) 표층 수온을 복원하는데 이용하는 것이 가능하다(Broecker, 1986; O'Brien et al., 2017, Fig. 1).
생성물(product)의 동위원소 비는 반응속도, 혹은 반응에 이용되는 기질(substrate)의 동위원소 비에 의해 결정되기도 한다. 해수 내 탄소를 이용해 에너지를 생산하는 식물플랑크톤은 대기와 해수 간 탄소 종에서 발생한 분별작용이 적용된 용존무기탄소를 이용하기 때문에 탄소의 동위원소 비가 달라지며, 식물의 광합성 반응 기작의 차이에 따라 나뉘는 C3 식물과 C4 식물은 탄소 고정 경로의 차이에 의해 다른 탄소동위원소 비를 가지게 된다. 이러한 차이는 상업적으로도 활용되기도 하는데 현재 벌꿀의 순수 여부 판단에 AOAC (Association of Official Analytical Chemists: Official methods of analysis)에 등재되어 통상적으로 이용되는 방법이 탄소 안정동위원소 분석법이다. 이는 식물의 탄수화물이 광합성 경로의 탄소 고정 방법에 따라 다른 것을 이용한 것으로 벌꿀을 생산하는 밀원이 대부분 C3 식물로 이루어져 있다는 것에 착안되었다(Cho et al., 2012). 즉, 사탕수수(C4 식물)에서 기인하는 설탕을 먹인(사양) 꿀을 동위원소 비의 차이로 구별하는 것이다. 이러한 접근은 과학수사에도 이용되는데, 2016년도 국내에서 발생한 일명 '농약소주' 사건은 소주에 포함되어있던 농약성분(methomyl)의 탄소, 질소 그리고 산소 안정동위원소 비를 분석해 이 값이 일치하는 농약성분의 동위원소 비로 범인을 확인했다. 이는 농약의 제조사에 따라 원료와 제조공정이 달라 제품의 동위원소 비가 다르다는 점을 이용한 것이다(Song et al., 2018).
생태학 분야에서 탄소와 질소의 안정동위원소 비 분석은 먹이망 구조(food web structure)와 섭식관계(feeding relation), 그리고 먹이망 내에서의 영양학적 지위(trophic position, TP)와 생태계 내에서의 영양학적 영역(e.g., isotopic ecological niche)과 같은 정보를 쉽게 파악할 수 있도록 도움을 주는 중요한 기법 중 하나다. 탄소의 안정동위원소 비 값이 섭식에 의한 분별작용에 의해 약 1‰ 정도의 증가를 보이기 때문으로 이 값을 이용해 생물의 에너지원(먹이원)을 추정할 수 있다. 반면 질소의 경우 양양학적 위치를 추정할 수 있는데, 이는 먹이생물과 섭식자 간 질소 대사에서 발생하는 분별작용으로 약 3.4‰의 일정한 증가가 관찰되기 때문이다(Minagawa and Wada, 1984, Fig. 2).
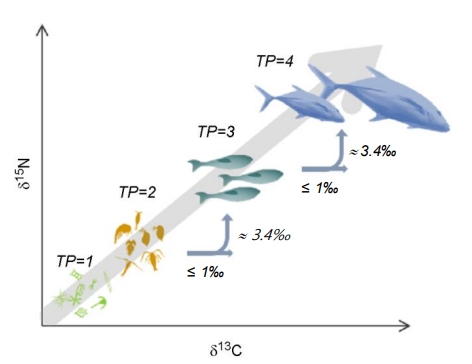
안정동위원소 비를 이용한 섭식관계와 먹이망 해석은 섭식에 대한 직접 관찰이나 위 내용물 분석(gut contents analysis)과 같은 조사 순간의 먹이원 보다 장기간에 걸쳐 축적된 정보를 반영한다는 점에서 야생동물과 어류의 생태계 내 영양학적 영역 조사에 매우 유용하다. 그리고 위 내용물 조사가 어려운 소형생물의 경우 적은 양의 시료로 동위원소 분석이 가능해 생태계 전반적 변화 양상을 조사 및 진단하는 연구가 가능하다(Polito et al., 2011; Ha et al., 2013). 나아가 안정동위원소를 이용한 먹이망 연구 결과는 생태계 모델 개발에 보다 정확한 생태학적 관계에 대한 정보를 제공할 수도 있는데, 섭식관계와 먹이의 기여도를 산정이 가능함을 보여주는 예가 mixing model(예, IsoSource)이다(Phillips et al., 2014). 이는 분별계수에 물질 수지 개념(mass balance)을 더해 먹이와 섭식자 관계와 먹이의 기여도를 해석하는데, 갯벌이나 심해, 극지 환경 등 다양한 생태계에서 에너지 흐름에 있어 구성 요소의 중요성을 해석하는 데에 유용하게 이용된다(Ha et al., 2013; Suh and Shin, 2013; Kang et al., 2016; Wang et al., 2018). Suh와 Shin의 연구는 대한민국 서해안 바지락 양식장에서 바지락 Ruditapes phillipinarum의 크기에 따른 먹이원의 변화를 해석 해 바지락의 성장에 따라 필요로 하는 양식장의 먹이 환경에 대해 토의하기도 했다(Suh and Shin, 2013).
안정동위원소 비 분석법을 이용한 환경 및 생태 연구는 1990년대 후반에 들어 Gas chromatography (GC)로 분리된 유기화합물에서의 안정동위원소 비 분석으로 발전하게 되었다. GC에 off-line 또는 on-line 연소 시스템으로 결합된 IRMS를 GC-IRMS라고 하는데, 이는 GC에서 분리된 화합물에서 동위원소(C, H, N) 분석이 가능하게 하여 환경유해물질(e.g., PAHs, pesticides)이나, 유기물을 구성하는 아미노산, 지방산과 같은 화합물을 대상으로 꾸준한 연구가 수행되고 있다(Larsen et al., 2009; Chikaraishi et al., 2014; Go et al., 2022a; Go et al., 2022b; Yun et al., 2022; Won et al., 2022). 이러한 분석 기술의 발전은 이전까지의 전체 시료(bulk tissue)를 이용해 분석하는 안정동위원소 비가 보여줄 수 있는 정보와 화합물이 가지고 있는 정보, 그리고 화합물의 대사 반응에서 관찰되는 정보까지 추가로 활용할 수 있다는 점에서 보다 많은 생리 생태적 해석이 가능하게 함으로써, 생태, 환경, 생물학 분야 등 다양한 분야에서 유망 기술로 주목 받고 있다(Choi et al., 2019; Won et al., 2020; Won et al., 2022; Yun et al., 2022, Fig. 3).
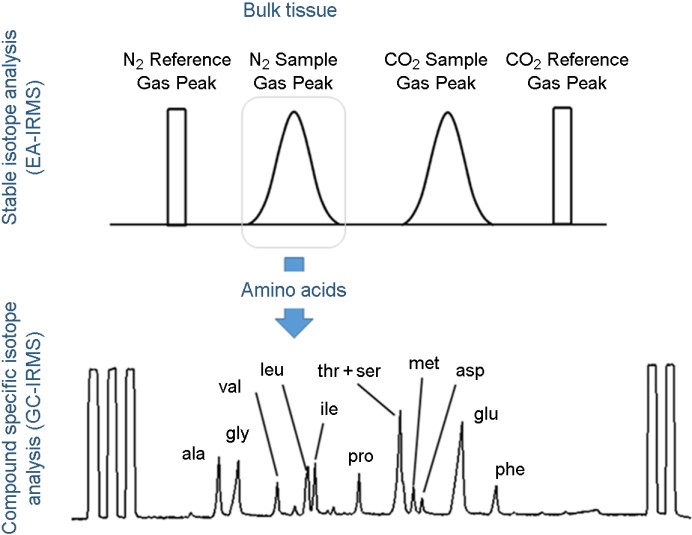
Fig. 3. IRMS connected with GC or LC can be used to measure isotope ratios of each compound. Upper and down pannel in- dicate nitrogen peak in bulk tissue measured by EA-IRMS and peaks of each amino acid separated from bulk nitrogen by GC-IRMS, respectively.
그 예로 아미노산을 대상으로 하는 탄소의 안정동위원소 비 연구는 동위원소 값의 유의미한 차이를 통해 먹이원을 추정에 유리하다. 그뿐만 아니라 끝단 값(end-member) 비교를 통한 finger- printing(특이적인 아미노산 동위원소 비의 패턴 분석법) 연구가 가능하게 하게 하는데, 섭식관계에서 체내 합성이 되지 않아 먹이로부터 전달되는 필수 아미노산의 탄소 안정동위원소 비로 박테리아, 미세조류, 그리고 육상식물로부터의 탄소 기원을 구별한다(Larsen et al., 2009). 또 다른 예로 아미노산의 질소 안정동위원소 비를 이용한 영양단계 산정은 기존 bulk tissue의 탄소와 질소 안정동위원소 비를 이용한 방법에 비해 정밀한 영양단계 계산이 가능하다(Chikaraishi et al., 2014). 이 방법은 생물의 체내 대사에 따라서 (1) 동위원소 비의 변화가 거의 없어 먹이(source, 에너지) 기원을 보여주는 아미노산(source amino acid)과, (2) 영양단계에 따라 개체 내 대사에 의해 일정한 비율로 그 값이 무거워지는 아미노산(trophic amino acid)의 동위원소 비의 차이를 이용하며, 하나의 시료(개체)에서 분석한 아미노산들의 질소 안정동위원소 비 결과로 보다 신뢰도 높은 영양단계를 계산할 수 있어 기존의 총 질소의 안정동위원소 비를 이용한 방법이 갖는 환경에 따른 변동성에 의한 불확실성과 생산자나 낮은 단계의 소비자를 포함하는 먹이생물을 함께 분석해야 했던 노력과 시간을 줄인다는 장점을 갖는다(Won et al., 2020).
최근 발표된 안정동위원소 분석 기법에 대한 리뷰는 위의 내용을 포함하는 생태학적, 지구화학적인 연구에 초점을 맞추고 있다(McMahon and McCarthy, 2016; Ishikawa et al., 2018; Ohkouchi et al., 2017; Whiteman et al., 2019). 본 총설에서는 안정동위원소 비 분석 기법을 적용하기 위한 이해를 목적으로 자연 값에 기반한 안정동위원소 비가 유용한 도구로 이용될 수 있는 이유(분별작용)를 설명하고 수산생물, 또는 양식생물을 대상으로 이용될 수 있는 환경, 생물 생리 및 생태 연구에 활용된 연구를 소개하고자 한다.
안정동위원소 비에 대한 이해: 분별작용과 표기(δ, ‰)
동위원소의 분별작용은 두 가지로 그 중 하나인 평형동위원소 분별작용(equilibrium fractionation)은 화학적 평형 상태(정반응 속도와 역반응 속도가 같아 겉보기 화학 반응이 일어나지 않는 것처럼 보이는 상태)에서 둘 이상의 물질 사이의 동위원소가 부분적으로 분별되는 것을 의미한다. 평형 분별작용은 더 무거운 동위원소가 기질이나 생성물과 더 강한 결합을 갖고 있어 화학 평형 반응의 기질과 생성물이 동위원소 비율이 다를 때 발생한다. 예를 들어, CO2와 H2O 분자 사이의 가역적인 산소 교환은 18O가 16O보다 탄소와의 더 강한 결합을 생성하기 때문에 18O가 많은 CO2를 생성하게 된다(Eq. 1).
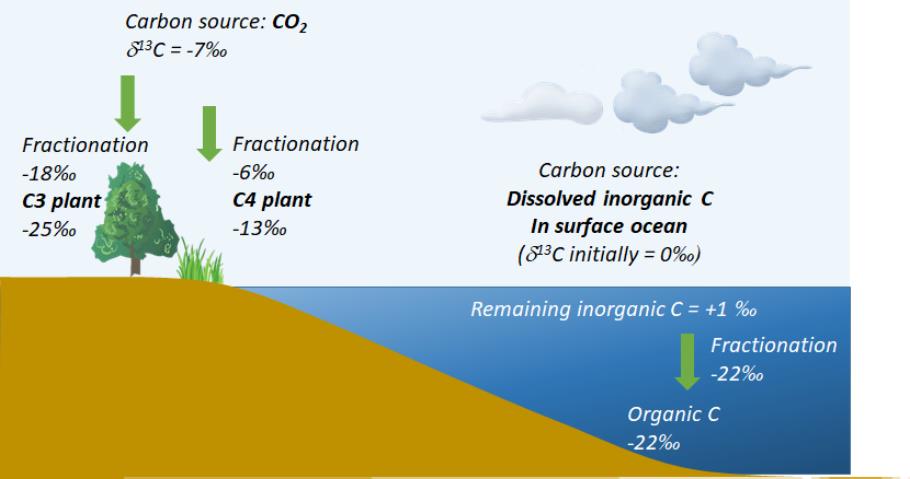
또 다른 반응으로 역학적 분별작용(Kinetic fractionation)은 분자가 상을 변경할 때(예: 액체에서 증기로) 또는 화학 반응이 비 가역적일 때 발생하며 한 방향의 반응에서 안정동위원소가 질량에 의해 분별되는 과정이다. 생물학적 과정이 "역학적" 동위원소 분별작용의 좋은 예가 되는데, 일반적으로 모든 유기체는 더 가벼운 동위원소를 우선적으로 사용하기 때문이다. 그 이유는 "에너지 비용"이 더 낮기 때문이며, 그 결과 기질(무거운 것)과 생물학적으로 생성된 생성물(가벼운 것) 사이에 질량 비의 차이, 다시 말해 분별(분획, fractionation)이 발생한다. 대표적인 예로 광합성작용에서는 대기 중 CO2 분자를 흡수하여 탄수화물을 합성하는 과정에서 12C의 가벼운 탄소동위원소를 우선적으로 이용하게 되며 이러한 반응 과정에서 동위원소의 분별로 인해 식물 및 식물의 탄소동위원소 비가 기질인 무기 탄소에 비해 가벼운 값을 가지게 된다(Fig. 4). 또한 이러한 반응은 무기 탄소를 이용하는 광합성 경로(기작)에 따라 다르기 때문에 Fig. 4에서와 같이 C3 식물과 C4 식물, 그리고 수중의 이산화 탄소를 이용하는 식물플랑크톤의 경우에 각각 다른 분별 값을 가지고 다른 탄소 안정동위원소 비를 나타내게 된다. 이와 같은 분별작용의 영향은 유기체의 질소에서도 관찰되는데, 이미 언급한 섭식관계에서 소비자에서 관찰되는 약 3.4‰에 해당하는 질소 안정동위원소 비의 분별작용은 소비자 생물 체내 질소의 대사에 의한 결과이다.
수산/해양 생물과 양식 환경에 적용한 안정동위원소 비 분석 연구 소개
1. 먹이질 및 형태 연구
생물을 대상으로 해양에서 수행되고 있는 많은 연구들은 특정 생물의 식이 특성이나 생태계 구성원 간 상호관계, 나아가 생태계의 구조를 이해하는 것에 초점을 두고 있다. 특히 생태계의 구성원 간 관계를 이해하기 위한 먹이-섭식관계나 먹이망 조사가 그 예가 될 수 있는데, 앞서 언급한 것과 같이 안정동위원소 비 분석 기법은 영양단계(TP) 해석을 위한 유용한 기법의 하나로 여겨지고 있다(Minagawa and Wada, 1984). 특히 화합물의 안정동위원소의 분석 기법이 발전하며 총 질소가 아닌 아미노산의 질소 안정동위원소 분석은 보다 정확한 영양단계 산정을 가능하게 했는데 이는 아미노산에 따라 영양단계에서 관찰되는 고유한 분별계수(TDF, trophic discrimination factor)의 차이를 이용한다(Chikaraishi et al., 2014; Won et al., 2020). TDF의 차이는 영양단계에 따라 큰 차이가 나지 않는 source amino acid와 그 차이가 크게 관찰되는 trophic amino acid으로 아미노산을 나눠볼 수 있으며, trophic amino acid으로 이용되는 대표적인 예인 glutamic acid의 δ15N는 영양단계에 따라 약 8.0‰ 정도 비교적 크게 증가한다(Fig. 5A). 이는 개체 내 glutamic acid의 대사 과정에서 아민기(NH3)기를 떨어뜨리는 과정에서 상대적으로 무거운 질소로 구성된 아미노산이 체내에 남게 되기 때문이다. 반면에 대표적인 source amino acid인 phenyl- alanine은 대사 과정에서 동위원소 비의 변화가 없으며 섭식을 통한 전이 과정에서도 거의 변화되지(0.4‰) 않아 먹이원에 대한 정보로 활용된다(Fig. 5A).
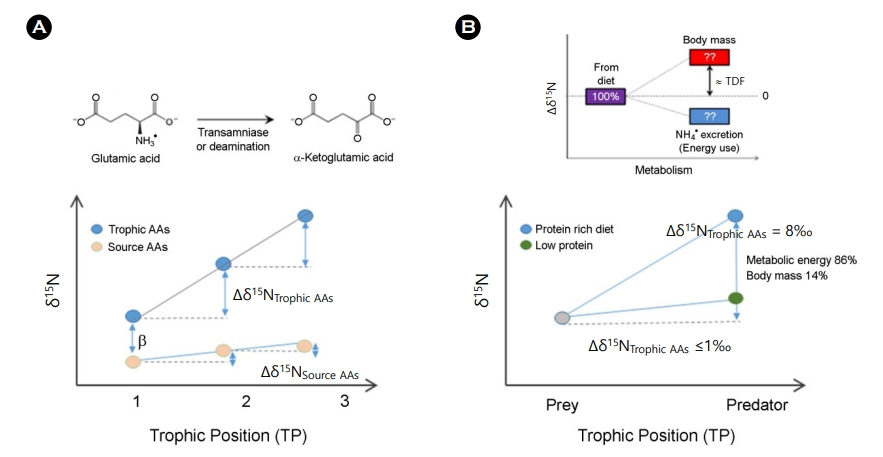
Fig. 5. Example of (A) trophic position study using compound specific isotope analysis of amino acid (nitrogen stable isotope ratio) and (B) study of TDF change in nitrogen stable isotope ratios of amino acid according to food quality (adopted from Miura and Goto, 2012).
나아가 Trophic amino acid는 생물의 체내 대사에 의한 변화를 나타낼 수 있는 아미노산인 만큼 먹이로부터 기인한 질소의 대사에 대한 결과도 보여줄 수 있음이 알려졌다. 먹이는 소비자가 에너지를 얻는 원천으로써, 먹이가 가진 영양소는 성장, 번식 그리고 그들의 생리적, 환경적 스트레스 요인 및 병원체에 대한 반응(저항) 등 건강성에 영향을 미치는 중요한 요소이다(Mzengereza et al., 2021). 야생의 생물을 대상으로 하는 연구에서 개체 또는 군집의 영양학적 건강성과 이들의 먹이 질을 파악하는 것은 영양학적 관계로 생태계의 기능적 면을 평가하는데 있어 중요한 요소가 되기도 하며 안정동위원소 비를 이용한 영양단계를 해석하는데 있어서도 고려해야 하는 사항이 된다(Chikaraishi et al., 2015). McMahon et al. (2015)은 단백질 함량과 아미노산 조성이 다른 먹이를 먹인 어류 mummichogs의 미성숙 개체를 이용한 실험에서 먹이의 종류에 따라 다르게 관찰되는 trophic amino acid의 TDF 변동성을 언급하며, 먹이가 가진 아미노산 조성과 섭식자가 필요로 하는 아미노산 조성의 유사성 정도와 단백질의 조성 등을 포함하는 먹이의 질의 다양성이 deamination 정도를 결정지어 아미노산의 질소 안정동위원소 비가 달라짐을 설명하고 있다. Chikaraishi et al. (2015)의 연구에서도 양서류 Bufo japonicus의 올챙이를 대상으로 한 먹이 급이 실험에서 공급한 먹이의 조성, 즉 단백질의 유무에 따라 glutamic acid를 포함하는 아미노산(alanine, valine, leucine, isoleucine, and glycine)의 TDF 값이 달라질 수 있음을 보여주었다. 단백질의 함량이 낮은 먹이 조건(탄수화물 기반 먹이)에서는 다른 조건(commercial pellet과 bloodworm을 급이 한 조건) 및 자연 생태계의 먹이 조건에서 일반적으로 관찰되는 8.0‰ 보다 작은 값의 분별(0.6‰)을 보였는데, 이 결과는 먹이로부터 얻을 수 있는 아미노산이 달라 발생하는 결과로 deamination의 플럭스의 차이 때문이다. Glutamic acid를 포함하는 일부 아미노산은 transamination과 deamination 과정을 통해 암모니아를 형성하는 과정을 통해 에너지를 생산하며 이론적으로 glutamic acid는 15N이 농축되는 동위원소 분별이 일어나 약 8.0‰의 TDF가 관찰된다고 알려져 있다. 그리고 이 8.0‰의 분별작용은 약 86%의 대사를 설명하며 나머지 14%가 미반응 상태로 남아 있음을 나타낸다(Miura and Goto, 2012) (Fig. 5B). 그러나 단백질이 부족한 먹이를 섭취한 경우 섭식자는 먹이에 기인한 아미노산으로부터의 에너지를 얻지 못하게 되기 때문에 먹이의 단백질(아미노산 pool)이 적어 설탕, 지질과 같은 다른 에너지원으로부터 대사에너지를 얻게 되기 때문에 유기체를 구성하는 아미노산은 최소한의 15N 농축만이 가능해 작은 TDF가 나타나며, 이는 낮은 대사율을 의미한다.
먹이의 원료 선택에 있어 생물의 먹이 이용성도 중요한데, 이는 양식생물의 생산성과 경제성에 있어 중요한 요소가 되기 때문이다(Mzengereza et al., 2021). 지금까지 동물성 단백질과 식물성 단백질을 비교하는 등의 먹이의 원료를 선별하기 위한 연구에서는 소화관련 효소나 유전자의 활성도를 분석하거나 개체의 성장이나 면역 능력을 비교하는 등의 연구가 진행되어왔다(Velazco-Vargas et al., 2014; Mzengereza et al., 2021). 최근 연구에서는 지질과 탄수화물의 대사의 차이를 측정하기 위해 아미노산의 탄소 안정동위원소 비를 생체지표로 활용할 수 있음이 보고되었다(Wang et al., 2019). 이 연구는 먹이에 따라 다른 소화율과 지질 대사 경로가 아미노산의 13C의 차이(먹이와 양식생물 간)를 야기한다고 설명하며 소화율이 높은 탄수화물과 낮은 소화율을 갖는 탄수화물 공급원, 즉 옥수수 전분과 macroalga (Palmaria palmate)를 먹은 Atlantic salmon에서 해당작용(glycolysis)에 관여하는 아미노산의 13C가 다른 결과를 보여주었다. 특히 지질의 탄소 안정동위원소 비가 가벼운 것을 고려하면 Palmaria를 급이한 Atlantic salmon에서 조직과 먹이 사이의 아미노산의 탄소 안정동위원소 비의 더 큰 차이는 에너지원으로서 먹이로부터 기인한 지질에 대한 수요가 증가함에 따라 비필수 아미노산(nonessential amino acids)의 대사 중간체를 생합성 하는 데 사용할 수 있는 지질의 비율이 감소됨을 설명한다. 또한, 두 그룹 간 필수아미노산(essential amino acids)의 유사한 13C 값을 이용해 모든 육상(완두콩 단백질 농축물 및 밀 글루텐 가루) 및 해양(어분 및 홍조류) 유래 단백질 공급원에 대해 유사한 영양소 동화 효율을 보여준다는 것을 해석하였다.
먹이의 질은 양식 산업이 집중해야 하는 필수적인 영역이라고 할 수 있다. 최근 다양한 수산생물이 양식 시스템에서 길러지고 있다. 야생에 서식하는 어류의 먹이를 대체하면서도 경제성을 갖는 고품질의 배합사료 생산은 고품질 양식 산업에서 중요한 연구 주제이다 안정동위원소 비를 이용한 먹이 연구는 양식생물의 영양관리에 도움을 줄 수 있으며, 이는 영양학적으로 흡수된 영양성분을 조성을 파악할 수 있게 하는 것에 그치지 않고 영양소를 활용하고 대사 중간체를 조직으로 보내는 방법에 대한 이해를 높여 나아가 경제성 있는 양식생물의 먹이 개발에 활용 될 수 있음을 보여준다.
2. 생리 및 습성 연구
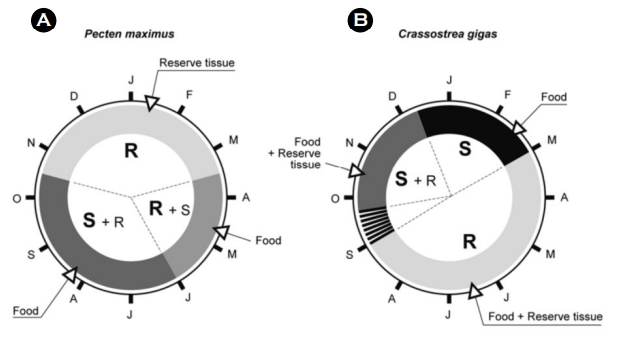
생물의 성장은 주요 생리적 반응 즉, 에너지 획득(섭식과 동화)과 에너지 소비(대사, 배설)의 결과로 나타나는데, 이러한 생리 과정에서 관찰되는 에너지 할당을 이해하면 생물의 건강성과 성장잠재력(scope for growth)을 추정할 수 있다(Paulet et al., 2006). 생물의 조직에서 관찰되는 안정동위원소 비의 차이는 이러한 에너지 할당에 대한 정보를 제공할 수 있는데 이는 조직에서 관찰되는 동위원소 값이 변하는 속도는 회전율과 성장을 포함한 대사 활동의 함수이기 때문이다. Paulet et al. (2006)의 연구는 이매패류 가리비 Pecten maximus와 굴 Crassostrea gigas에서 먹이로부터의 흡수된 탄소가 체내 반영되는 정도를 통해 에너지 할당 패턴을 확인하고자 하였다. 이를 위해 기존에 섭식하던 먹이와 다른 동위원소 비를 갖는 새로운 먹이(depleted 13C)을 제공한 후 여러 기관(gonad, adductor muscle, digestive gland 그리고 gills)의 동위원소 비를 분석한 결과 점차적으로 먹이의 동위원소 값이 반영되어 그 값이 변화하는 것을 확인할 수 있었고, 또한 기관별, 계절 및 종간 차이가 있음을 확인하였다(Paulet et al., 2006). 해당 연구에서 먹이에 기반한 에너지는 조직에 반영된 안정동위원소 비의 변화에 이용해 carbon incorporation index (CII)로 표기되었는데, digestive gland 에서 관찰된 가장 빠른 CII와 adductor muscle의 가장 느린 CII는 먹이로부터 획득한 에너지의 대사 활성이 조직에 반영되는 차이를 보여주었으며, 굴보다 높은 가리비의 CII는 먹이로부터 획득한 에너지의 대사 활성도의 종 간 차이가 있음을 보여주었다. CII의 계절적 차이도 관찰되었는데, 이러한 해석은 계절에 따른 패류의 에너지 할당 시나리오를 보여준다. 이미 보고된 바에 의하면 가리비의 경우 봄과 여름에는 체질량 유지와 생식에 필요한 에너지를 섭식을 통해 직접 공급받으며, 겨울철에는 봄과 여름 동안 저장된 에너지를 생식과 체질량(body mass) 유지에 사용하고 이는 보통 adductor muscle에 저장된 글리코겐과 digestive 저장된 지질인데(Fig. 6, Paulet et al., 2006), 본 연구는 이 같은 이전의 시나리오를 뒷받침 하면서도 안정동위원소 비를 이용한 해석을 통해 이 종의 겨울에 관찰되는 생식선 활동이 저장 조직에 직접적으로 의존한다는 사실과 기존의 시나리오 보다 빠른 봄 철 섭식 가능성에 대한 해석까지 강조하고 있다. 이 같은 시도는 종에 따라 다른 계절적 에너지 할당에 대해 계절적 주기를 바탕으로 한 번식 생리 해석에 활용될 수 있으며 나아가 양식 환경의 최적화를 위한 유용한 정보가 될 수 있음에 따라 안정동위원소 비를 이용한 생물의 대사평가의 응용 가치를 확인시켜준다.
최근 연구에서는 이 같은 동위원소 비의 변화로 먹이로부터 탄소 동화가 제한된 상태에서 일어나는 에너지 이용에 보여주기도 하였다. 35일 간 수행된 절식(fasting) 실험에서 방어과의 amber- jack Seriola lalandi는 체내 crude lipid를 에너지원으로 가장 많이 사용하는 것으로 나타났으며 반면 다가불포화지방산(Polyunsatu- rated fatty acids, PUFA) 및 long chain의 PUFA는 보존되는 결과를 보였다(Barreto-Curiel et al., 2017). 안정동위원소 비 분석을 이용한 해석에서는 근육과 간을 비교하였는데, 특히 δ15N 및 아미노산의 탄소와 질소의 안정동위원소 비(δ13C-AAs 및 δ15N-AAs)는 근육에서 차이가 없었던 반면, 간에서는 그 값이 증가하는 결과를 보였으며 δ13C 및 δ15N의 증가는 단백질 turnover가 발생했음을 나타내 절식 기간 동안 에너지 생산 과정으로의 신진 대사 경로를 위한 소비에 대한 증거가 되었다. 이처럼 특정 조직에서 관찰되는 대사의 변화에 대한 지표로서의 안정동위원소 비의 활용은 수산생명의 질병학이나 수산품질평가 등 다양한 분야에서의 접근 역시 가능할 것을 예측하게 한다.
그 밖에, 아미노산 안정동위원소 비를 이용해 야생의 동물에 대한 비파괴적이면서도 장기간 누적된 먹이를 해석하는 연구에 활용되기도 한다. Gómez et al. (2018)은 이동하는 동안 철새의 먹이를 해석하기 위해 철새 gray-cheeked thrush의 다양한 조직과 과일 및 곤충 등 먹이에서 아미노산 탄소 질소 안정동위원소 비를 분석했다. 그 결과 깃털과 발톱에 비해 혈액의 영양학적 위치가 낮고, 중간 기착 일수에 따른 혈액 내 source amino acid인 phenylalanine의 δ15N 값이 감소한 결과를 확인하였으며, 이는 이들이 겨울철 이동하는 동안 번식기나 월동기 보다 더 많은 과일을 소비했음을 설명한다. 이 연구는 아미노산의 동위원소 분석이 철새의 먹이 망, 계절별 식이 변화 및 영양소의 대사 경로를 이해하는 데 어떻게 기여할 수 있는지 보여주는 좋은 예가 되었다. 유사한 연구로 우리나라에서는 회유성 어종인 뱀장어를 대상으로 하는 안정동위원소 비 분석 연구를 이용해 이동 중 섭식에 대한 습성을 해석하기도 하였다(Kim et al., 2016). 우리나라에서 발견된 실뱀장어는 마리아나 해구에서 산란, 부화된 후 북적도 해류를 따라 회유하는 어종으로 낙동강 하구에서 채집된 개체의 탄소 및 질소 안정동위원소 비는 북적도 해류에서 채집된 뱀장어 자어인 렙토세팔루스(leptocephalus)와 유사한 범위를 보였으며, 이러한 결과는 이 종이 회유 기간 동안 먹이 섭취를 하지 않는 결과로 추론되었다(Kim et al., 2016). 이는 이전에 조사된 연구에서 뱀장어의 자어와 입자유기물질과의 안정동위원소 증가가 일치하는데 반해 낙동강 하구에서 조사된 입자성 유기물질의 안정동위원소 비의 값이 실뱀장어의 값과는 매우 큰 차이를 보여 먹이 기원으로서의 가능성이 낮게 판단되었기 때문이다. 특히, 강의 입구에서 발견된 실뱀장어의 공복률과 복강의 지질을 이용한 에너지 이용의 가능성(Kawakami et al., 1999), 그리고 실내 실험에서 인공사료의 공급에도 불구하고 섭식이 관찰되지 않았던 점 등이 이러한 해석을 뒷받침한다고 설명하고 있으며, 이는 우리나라에서 양식의 종묘로 이용되는 자연산 실뱀장어에 대한 이해를 높여 관련 산업의 활성화뿐만 아니라 인공 사육 시스템 등의 사육 환경 조성에 중요한 정보 제공의 결과를 보여준다.
3. 양식 환경 연구
환경에 존재하는 유기물의 안정동위원소 비의 차이는 유기물이 생성되는 과정(기작)과 환경을 구분하는 지표로 이용할 수 있다. 대기의 탄소와 수중의 HCO3를 이용하는 1차 생산자의 다른 탄소 안정동위원소 비의 값이나 가축 분뇨와 합성비료에서 기인한 질소의 안정동위원소 비가 구별되는 것은 유기물이 생성되는 당시의 원료과 생성 기작의 차이를 보여주며, 해양생물의 경우 고위도에 위치할 수록 가벼운 탄소 값을 갖는 등 반응이 일어난 환경 역시 동위원소 비에 반영될 수 있다. 질소의 경우도 마찬가지인데, 대기 중의 질소는 0‰의 값을 나타내며, 식물에 의한 생물학적 질소 고정은 그 분별작용이 작아 식물의 질소동위원소의 값은 대기와 거의 같은 값을 갖는다. 그러나 해양기원 유기물은 이보다 무거운 값을 가진다고 알려져 있으며, 가축 분뇨에서 발생한 암모니아 화합물로 만들어진 합성비료는 이보다 20~30‰ 가량 무거운 값을 나타낸다. 이러한 이유로 안정동위원소 비는 유기물의 기원을 추적하는 데에 활용되며 환경 오염을 유발하는 유기물의 기원이나 생태계에서의 기저 에너지(먹이)를 판별하는 데에 활용되고 있다.
국내 양식이 이루어지고 있는 환경에서도 이를 이용한 유기물 기원 연구가 수행되었다. 양식장 내 환경 적합성은 다양한 요소에 의해 평가되고 이용되는데 그 중 양식생물의 배설물 및 섭취되지 못하고 잔류하는 먹이가 침전되어 발생하는 유기물 오염은 양식장 내 주된 오염 원으로 여겨지고 있다. 최근 어장의 회복력을 진단하기 위한 연구에 탄소와 질소의 안정동위원소 비가 활용된 바 있는데, 어장 내 퇴적 유기물의 기원을 해석해 유기물의 기여율을 계산하는 방법을 이용했다(Go et al., 2022a). 이 연구에서 이용하고 있는 Bayesian 혼합(mixing) 모델은 다수의 오염 원에 대한 기여율을 통계적 신뢰구간과 함께 제공하여 기여율 추정 결과에 대한 신뢰성을 향상시킬 수 있으며, 분별작용에서 기인한 불확실성을 수치로 보정하는 것이 가능하다(Nam et al., 2019). 이를 이용해 Go et al. (2022a)은 경상남도 하동군 중평항 숭어 가두리 양식장이 밀집된 연안 환경에서 과거 양식장으로 이용된 지점과 양식장의 영향을 받지 않는 대조군 지점에서 입자물질과 퇴적물 내 유기물 시료를 대상으로 유기 탄소 및 질소의 농도를 분석하였다. 그리고 다양한 기원의 유기물[C3 식물, 담수 기원 입자성 유기물질, 어류의 분변, 그리고 자생기원 유기물(식물성플랑크톤)]로부터 확인된 안정동위원소 비 끝단 값을 이용하여 퇴적유기물 기여도 산정과 절대 값 비교를 수행하였다. 그 결과 현재 어업이 이루어지지 않는 지점에서도 어류의 배설물과 자생기원 유기물의 높은 기여율이 관찰되었고, 이는 양식 기간 동안 퇴적된 양식 활동에 기인한 유기물이 양식 휴업 기간 동안에도 분해되지 않고 장기간 존재함을 의미했다. 또한 안정동위원소 비의 비교를 통해 양식장의 환경도 평가할 수 있었는데, 이는 먹이가 생물에게 흡수되어 배출되는 단계에서 발생하는 분별작용의 결과 먹이와 분해된 먹이, 그리고 소화된 형태의 배설물의 값이 다르기 때문으로 Yokoyama et al. (2010)과 Go et al. (2022a)는 양식어류의 배설물(feces)이 먹이(pellet diet)에 비해 가벼운 탄소 및 질소 안정동위원소 비를 보임을 보여주었다. 특히 Yokoyama et al.의 연구에서는 이러한 안정동위원소 비를 이용해 미쳐 섭식되지 못한 먹이의 양이 어류의 배설물로부터 기인한 유기물을 두 배 이상 초과하는 것을 확인하였으며 이러한 결과를 바탕으로 양식장에서 사료 과잉 공급 여부를 판단할 수 있음을 확인하였고 먹이의 급이 조건을 조절한 경우의 성장을 함께 비교하였다. 이는 어류의 성장과 폐사, 그리고 퇴적되는 유기물의 비교 연구를 통해 수산물에 이용되기에 충분하면서도 환경 수용 능력을 초과하지 않는 경제적이면서 어장의 지속 가능성을 개선하는 사료 공급 방식과 최적화 및 사료로 인한 환경 오염을 줄일 수 있는 연구가 가능함을 보여주었다(Yokoyama et al., 2010; Go et al., 2022a).
환경 시료 외에 생물을 구성하는 유기물의 안정동위원소 비 분석은 생물이 섭식한 유기물 에너지원에 대한 상대적인 기여도 계산 역시 가능하게 한다. Deudero et al. (2011)은 안정동위원소 비를 이용한 Bayesian mixing 모델을 기반으로 연안지역에서 양식 활동의 영향을 줄이기 위해 사용되는 생태통합양식법의 효율성을 평가하기도 했다. 생태통합양식법(IMTA, integrated multi-trophic aquaculture)은 양식 환경에서 발생되는 입자성 유기물의 저감을 위한 방안으로 활용되는 양식 방법으로 영양이용단계가 다른 양식생물을 일정 해역에 통합해 수산물을 기름으로써 배설물과 잔여먹이와 같은 입자성 유기물과 무기물을 활용하는 친환경 양식 기술로 각광받고 있다. Deudero et al. (2011)의 연구에서는 안정동위원소 비를 이용한 유기물 기여율 계산 결과 주요 양식 대상인 어류 Argyrosomus regius가 사료(pellet)와 fresh food에 각각 80.43% 그리고 19.57% 의존하고 있음을 확인하였고, 함께 양식된 scallop Chlamys varia의 경우 유기물의 약 70.03%에서 최대 93.81%가 pellet food를 비롯한 양식장으로부터 기인하는 물질(fresh food와 어류의 배설물)에 기인하고 있음을 확인하였다. 안정동위원소 비의 경향을 보더라도 양식장과 대조군 환경에서 배양한 두 종의 패류 scallop Chlamys varia과 mussel Mytilus gallo- provincialis는 양식장 환경에서 양식된 개체에서 분석된 풍부한 13C와 15N (enriched 13C와 15N) 결과를 보여 이들 이매패류가 효율적으로 양식장으로부터 기인한 물질을 동화하고 있음을 보여주었다.
이 외에도 안정동위원소 비를 이용한 먹이 기여도 평가는 생태계 구조에 있어 기저먹이원의 중요성을 해석하는데 활용되어 수산 양식 분야에 접목될 수 있다. 남해의 앵강만, 동대만, 그리고 광양만에 서식하는 어류군집을 대상으로 한 유기물 기여도 연구에서, Kang et al. (2009)은 하천의 어류군집이 육지로부터 기인한 갈대와 같은 습지생산자와 유사한 가벼운 값의 범위를 보이는 반면 상대적으로 높은 δ13C 값을 가지는 연안의 어류가 연안의 저서생산자가 가진 δ13C 값과 일치하는 범위를 보이며 연안에 서식하는 어류가 만 자체의 자생기원 저서생산에서 유래한 탄소원에 주로 의존하고 있다는 것을 보여주었다(Fig. 7). Kang et al. (2016) 역시 바지락 양식장이 위치한 영흥도의 조간대 갯벌에서 입자성 유기물과 저서미세조류, 퇴적물 내 유기물과 해조류 및 잘피(Zostera marina)의 탄소 및 질소 안정동위원소 비를 분석하여 이매패류, 갑각류, 복족류, 어류를 포함하는 저서생태계와 비교함으로써 저서 생태계를 구성하는 가장 기초적인 탄소원이 저서미세조류(benthic micro algae)임을 보여주었다. 특히 이 연구에서는 저서 생태계를 구성하는 생물군의 탄소와 질소 안정동위원소 비의 차이를 통해 구분되는 세 개의 그룹에서 생태적 지위의 유사성을 해석해 이들 생활 및 섭식 특성에 기반한 생태계 구조를 해석하기도 하였다.

이렇듯 관심 수산 어종의 잠재적인 먹이생물까지 포함하는 접근은 자연 환경을 어장으로 하는 지역에서 자원생물의 서식지로서의 적합성을 평가하는데 활용될 수 있으며, 나아가 양식 및 수산생물의 생산성 연구에 중요한 자료로 활용될 수 있다. Suh and Shin (2013)에 의해 수행된 바지락 먹이원 해석 연구는 바지락의 성장에 따른 먹이원의 변화가 이들의 성장과 중요한 관련이 있음을 해석하기도 하여 생리변화에 따라 달리 요구될 수 있는 먹이생물과 서식 환경에 대한 토의에 안정동위원소 비 분석이 유용하게 활용될 수 있음을 보여주기도 하였다.
4. 수산물의 산지 판별(authentication)연구
"Isoscape"는 동위원소(isotope)와 영역(landscape)을 의미하는 단어에서 파생된 용어로 동위원소 분포의 지질지도(geologic map)를 의미하며 나아가 지리 정보 시스템(GIS)에서 동위원소 비율(δ)의 공간적이면서 명시적인 예측 모델(predictive model)의 의미를 포함한다(Bowen et al., 2009). 수소, 탄소, 산소, 질소, 스트론튬이나 황 동위원소 비는 유기체의 기원, 분포(또는 분배) 그리고 출처에 관한 과학적인 정보를 제공하는데, 이를 이용해 법의학적 감식이나 생지화학적 순환, 고고학, 오염 추적 등에 사용한다(Choy, 2018; Song et al., 2018; Hong et al., 2019). 이 방법은 작물이나 생물의 물 사용, 또는 섭식 활동이 환경 고유의 값을 반영하기 때문인데, 따라서 동위원소 비를 이용해 생물의 이동에 대한 정보를 해석하는데 이용되기도 한다. 그 예로 물의 수소 안정동위원소 비는 지형학적 특성과의 밀접한 관련 때문에(latitude, altitude, continental effect) 수문학적인 물의 순환 해석과 작물이나 과실에 반영된 값을 이용한 원료의 산지를 판별하는 정보로 이용될 수 있다. 그 예로 Gimenez et al. (1999)은 스페인의 여러 지역에서 생산된 포도에 대한 탄소와 수소 안정동위원소 연구 결과를 이용해 와인의 산지 판별에 대한 결과를 제시한 바 있으며, 그 밖에도 조류가 섭취하는 물이 깃털의 동위원소 비에 반영됨을 이용해 북미지역에서 철새의 이동 경로 연구에 활용되기도 하였다(Chamberlain et al., 1997; Hobson and Wassenaar, 1997). 어류를 대상으로 하는 연구에도 이러한 접근이 가능한데, 어류를 선택하여 이동 중 일시적으로 머문 기간과 지리적 정보를 연결하고 집단을 동위원소로 특성화 시켜 동위원소의 유사성에 근거한 지리적인 위치를 추론하여 집단과의 연관성을 찾는 방법이 그것이다. 실제로 어류의 경우 이석(耳石, otolith)을 구성하고 있는 원소(탄소, 질소, 산소, 스트론튬 등)는 바다, 하구, 강 등 물의 동위원소 비를 반영하기 때문에 이를 분석함으로써 어류의 이동 경로를 추정할 수도 있다(Trueman et al., 2012).
최근에는 이러한 동위원소 비의 특성을 이용하여 수산물의 양식 여부나 산지를 구별하는 데에 활용되고 있다(Thomas et al., 2008). 여러 환경 문제가 발생하는 지금 시대에는 식료품의 산지를 판별하는 것은 식품의 품질, 그리고 안전성과 연관되며 이는 소비자가 상품을 선택하는 데에 중요한 요소가 된다. 우리나라 역시 먹거리에 대한 안전과 소비자의 정보 요구를 보장하기 위해 생산지에 대한 표기를 의무화하고 유통 경로 추적과 같은 관리를 위한 시스템이 제시되고 있으며, 유럽의 경우에도 2002년 어류와 어류를 이용한 가공품에 대해 양식 여부에 대한 정보를 표기하게 하는 등 노력이 있어왔다. 그러나 수산물의 경우 가공품과 달리 자연에서 산물이 채집된다는 점에서 위조가 발생하기 쉽고, 이렇게 발생할 수 있는 사기행각을 줄이기 위한 노력으로 다양한 연구가 수행되고 있다(Won et al., 2021). 수산물을 대상으로는 형태학적 비교, DNA 염기서열을 통한 종이나 군집 구별, NMR을 활용한 분자 연구 등 여러 기법이 시도되고 있으며, 안정동위원소 분석 기법은 그 중 서식지에 대한 정보를 통한 산지 및 양식법 판별 기술로 각광받고 있다.
Thomas et al. (2008)은 자연산과 양식이 된 연어 Salmo salar 를 구분하는 데에 안정동위원소를 이용했는데, 이는 자연산 물고기와 양식 물고기 사이에 먹이의 구성의 차이가 조직을 구성하는 화합물에 반영되는 것을 이용한 것이다. 저자는 유럽, 북미 및 테즈메이니아 내 32개 산지에서 채집된 자연산 및 양식 언어 171개체를 대상으로 안정동위원소 비를 분석했고 그 결과 분석에 이용한 개체의 부분 시료(지질, 콜린, 글리세롤, 지방산, 근육, 수분) 중 choline에서 측정한 질소 안정동위원소 비(δ15N)와 total oil에서 측정한 산소 안정동위원소 비(δ18O)가 뚜렷한 차이를 보이며 이들의 서식 환경(자연산 개체, 양식 개체, 그리고 양식장에서 도망 나온 개체)을 구별해 주는 것을 확인했다.
최근에는 변별력을 높이고자, 3종 이상의 원소를 이용한 동위원소 기법을 이용한 다중 동위원소 분석을 통해 산지 판별을 시도하고 있다. 우리나라에서는 탄소와 질소, 그리고 황의 안정동위원소 비를 이용해 바지락의 원산지 판별과 관련된 분석 기법에 관한 특허와 논문이 발표된 바 있는데, 이는 기존의 유전자 정보를 가지고 분석하던 산지 판별법이 가진 한계를 보완할 수 있다. 우리나라에서 생산되는 바지락의 경우 중국에서 수입된 치패를 이용하기도 하는데 실제로 우리나라는 연 4,000톤의 치패를 중국으로부터 수입해 국내 양식에 이용하고 있어, 2009년 이후부터는 이미 유전형을 이용한 바지락의 산지 판별이 불가능한 실정이다(KIMST, 2017; MOF, 2020). 따라서 바지락이 서식한 지역의 정보를 반영할 수 있는 지표를 이용한 산지 판별로서 안정동위원소 기법이 제시되었는데, 우리나라와 중국의 해수와 퇴적물에서 관찰되는 동위원소 비의 지역적 차이가 바지락에 반영되어 이를 이용한 구별을 시도한 연구이다(Won et al., 2021). 저자는 바지락의 adductor muscle에서 탄소(δ13C), 질소(δ15N), 산소(δ18O), 수소(δD), 황(δ34S)을 비롯해 스트론튬(87Sr/86Sr)과 네오디움(εNd)의 안정동위원소 비를 분석하여 이들 원소의 교차 적용 및 선형판별분석법(LDA)을 이용해 생산지 판별에 최대 판별력을 갖는 원소 조합을 찾고자 하였으며, 그 결과 유기물의 기원(탄소)과 영양단계 및 배경질소를 포함하는 먹이망 정보(질소), 지질학적인 정보(황 과 스트론튬)를 이용한 조합에서 북한산, 중국산, 그리고 국내산을 구별할 수 있음을 보여주었다(Won et al., 2021, Fig. 8).
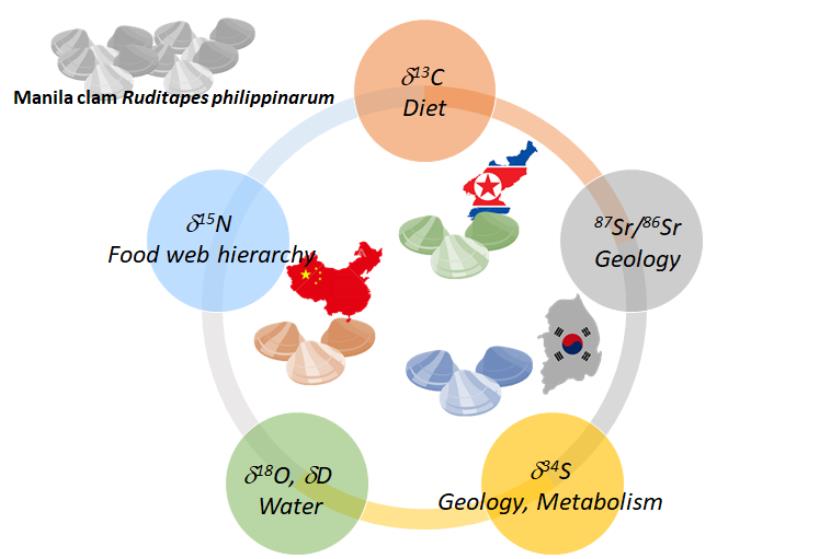
이후 바지락의 먹이가 되는 일차생산자의 정보를 보여줄 수 있는 바지락의 지방산 조성과 지방산을 구성하는 탄소의 안정동위원소 비(δ13C-FA)의 추가한 연구도 수행되기도 했다(Go et al., 2022b). 지방산의 조성과 지방산의 탄소 안정동위원소 비는 해수 중 바지락의 먹이가 될 수 있는 부유성 유기물(식물플랑크톤과 입자성 유기물질 등)의 정보를 포함하고 있어 기존의 조직 내 탄소와 질소, 황 등의 다원소 분석법에 기초한 기존의 산지 판별 시도와 비교해, 수입 바지락에 대한 산지 뿐만 아니라 국내의 네 개 연안(충청도, 전라북도, 전라남도, 그리고 경상남도)을 구별하는 등 강력한 판별력을 보여주기도 하였다(Go et al., 2022b).
향후 연구방향
안정동위원소 분석 기법은 유기물의 기원을 추적하고, 육상 및 수 생태계에서 먹이망의 구조를 밝히거나 동물의 이동 경로를 규명하는 등 다양한 분야에서 적용되고 있다. 선진 외국에서는 1950년대 초반부터 다양한 연구 분야에서 안정동위원소 비를 활용한 연구가 진행되고 있으며, 국내에서는 최근에야 몇몇 연구기관에 기기가 마련되어 그 활용의 폭이 넓어지고 있는 실정이다. 특히 화합물의 안정동위원소 비 분석에 활용되는 GC-IRMS의 경우 국립환경과학원, 한국기초과학지원연구원, 국립과학수사연구소, 한국해양과학기술원, 한양대학교 등 기관에서 기기를 보유하고 연구를 진행 중으로 다양한 시료를 대상으로 한 결과가 축적되는 중이다. 본 총설에서는 해양생물의 먹이, 생리, 서식 환경과 생물 간 관계 등의 주제를 바탕으로 수산 · 양식 생물에 활용될 수 있는 연구의 내용과 결과를 소개하고 있다. 본문에서 언급한 바와 같이 안정동위원소 분석 기법은 다양한 분야에서 새로운 해석을 하거나, 기존의 이론을 견고하게 할 과학적 근거를 제시하는 데에 이용될 수 있으며, 환경과 생명이 가진 시공간적 차이에서 발생하는 차이를 해석하기 위한 생물학적, 생리학적, 환경적 해석 연구에 유용하게 활용될 수 있는 기법이다.
수산 및 양식 분야의 발전은 기후 변화에 직면한 우리 인간에게 식량자원 확보와 건강한 수 생태계 보전의 의무로 해석될 수 있다. 양식 · 수산 생물의 건강과 관련된 먹이, 생리, 성장, 번식과 같은 생물에 대한 정보 및 이들의 건강한 서식 환경과 유통에 이르기 까지 구축되어야 할 양식 수산 학의 분야의 환경은 안정동위원소 분석법의 새로운 적용의 장이 될 수 있을 것으로 기대한다. 추후 국내 수산 양식 분야의 연구에서 안정동위원소 분석법을 적용할 수 있는 주제를 발굴하고 적극 활용하여 안정동위원소 분야가 수산 및 양식 분야 연구 영역으로 확장 될 것을 기대한다.
- References
-
1. Barreto-Curiel F, Focken U, D'Abramo LR, Viana MT. 2017. Metab- olism of Seriola lalandi during Starvation as Revealed by Fatty Acid Analysis and Compound-Specific Analysis of Stable Isotopes within Amino Acids. PLoS ONE 12: e0170124.
-
2. Ben-David M, Flaherty EA. 2012. Stable isotopes in mammalian research: a beginner's guide. J Mammal 93: 312-328.
-
3. Bowen GJ, West JB, Vaughn BH, Dawson TE, Ehleringer JR, Fogel ML, Hobson K, Hoogewerff J, Kendal C, Lai C-T, Miller CC, Noone D, Schwarcz H, Still CJ. 2009. Isoscapes to address large-scale earth science challenges. EOS 90: 109-116.
-
4. Broecker WS. 1986. Oxygen isotope constraints on surface ocean temperatures. Quat Res 26: 121-134.
-
5. Chamberlain CP, Blum JD, Holmes RT, Feng X, Sherry TW, Graves GR. 1997. The use of isotope tracers for identifying popu- lations of migratory birds. Oecologia 109: 132-141.
-
6. Chikaraishi Y, Steffan SA, Ogawa NO, Ishikawa NF, Sasaki Y, Tsuchiya M, Ohkouchi N. 2014. High-resolution food webs based on nitrogen isotopic composition of amino acids. Ecol Evol 4: 2423-2449.
-
7. Chikaraishi Y, Steffan SA, Takano Y, Ohkouchi N. 2015. Diet quality influences isotopic discrimination among amino acids in an aquatic vertebrate. Ecol Evol 5: 2048-2059.
-
8. Cho Y-J, Kim J-Y, Kim M-I, Kang K-M, Park Y-C, Kang I, Do J-A, Kwon K, Oh J-H. 2012. A study on stable isotope ratio of circulated honey in Korea. Korean J Food Sci Technol 44: 401-410.
-
9. Choi B, Choi H, Shin K-H. 2019. Determination of nitrogen isotopic baseline using nitrogen isotope ration of individual amino acids in coastal environment. Sci Fish Oceanogr 28: 98-105.
-
10. Choy K. 2018. Sulphur isotope analysis of bone collagen from archaeological human remains in southeastern Korea: Inter- preting human migrations and paleodiets in Korean pre- history. Hanguk Kogo-Hakbo, 109: 40-65. (in Korean)
-
11. Deudero S, Tor A, Alomar C, Valencia JM, Sarriera P, Blanco A. 2011. Integrated multitrophic aquaculture: filter feeders bivalves as efficient reducers of wastes derived from coastal aquaculture assessed with stable isotope analyses, aquaculture and the environment - A Shared Destiny, Dr. Barbara Sladonja (Ed.), ISBN: 978-953-307-749-9, InTech.
-
12. Gilbert A, Robins RJ, Remaud GS, Tcherkez GGB. 2012. Intramo- lecular 13C pattern in hexoses from autotrophic and hetero- trophic C3 plant tissues. PNAS 109: 18204-18209.
-
13. Gimenez M, Salazar D, Solana I. 1999. Regional origin assignment of red wines from Valencia (Spain) by 2H NMR and 13C IRMS stable isotope analysis of fermentative ethanol. J Agric Food Chem 47: 2645-2652.
-
14. Go YS, Lee D-I, Kim C-S, Sim BR, Kim HC, Lee W, Lee D-H. 2022a. Origin and source appointment of sedimentary organic matter in marine fish cage farm using carbon and nitrogen stable isotopes. Korean J Ecol Environ 55: 99-110.
-
15. Go YS, Won E-J, Kim SH, Lee DH, Kang JH, Shin K-H. 2022b. Step- wise approach for tracing the geographical origins of the manila clam Ruditapes philippinarum using dual-element iso- topes and carbon isotope of fatty acids. Foods 11: 1965.
-
16. Gómez C, Larsen T, Popp B. 2018. Assessing seasonal changes in animal diets with stable-isotope analysis of amino acids: a migratory boreal songbird switches diet over its annual cycle. Oecologia 187: 1-13.
-
17. Ha S-Y, Min W-K, Kim D-S, Shin K-H. 2013. Trophic importance of meiofauna to polychaete in a seagrass (Zostera marina) bed as traced by stable isotope. J Mar Biol Assoc U K 94: 121-127.
-
18. Hobson KA, Wassenaar LI. 1997. Linking breeding and wintering grounds of Neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers. Oecologia 109: 142-148.
-
19. Hong S, Lee Y, Yoon SJ, Lee J, Kang S, Won E-J, Hur J, Khim JS, Shin KH. 2019. Carbon and nitrogen stable isotope signatures linked to anthropogenic toxic substances pollution in a highly industrialized area of South Korea. Mar Pollut Bull 144: 152-159.
-
20. Ishikawa NF, Chikaraishi Y, Takano Y, Sasaki Y, Takizawa Y, Tsuchiya M, Tayasu LI, Nagata T, Ohkouchi N. 2018. A new analytical method for determination of the nitrogen isotopic com-position of methionine: Its application to aquatic ecosystems with mixed resources. Limnol Oceanogr-Meth 16: 607-620.
-
21. Kang C-K, Choy EJ, Kim Y-S, Park HJ. 2009. δ13C evidence for the importance of local benthic producers to fish nutrition in the inner bay systems in the Southern coast of Korea. J Korean Soc Oceanogr 14: 56-52.
-
22. Kang S, Choi B, Han Y, Shin K-H. 2016. Ecological importance of benthic microalgae in the intertidal mud flat of Yeongheung Island; Application of stable isotope analysis (SIA). Korean J Ecol Environ 49: 80-88.
-
23. Kawakami Y, Mochioka N, Kimura R, Nakazono A. 1999. Seasonal changes of the RNA/DNA ratio, size and lipid contents and immigration adaptability of Japanese glass-eels, Anguila japonica, collected in northern Kyushu, Japan. J Exp Mar Biol Ecol 238: 1-19.
-
24. Kim JB, Lee W-C, Kim HC, Hong S, Park KD. 2016. Feeding habits of the glass eel Anguilla japonica determined by C and N stable isotopes in the Nakdong River estuary of the Korean Peninsula. J Korean Soc Oceanogr 21: 36-43.
-
25. KIMST, Korea Institute of Marine Science and Technology Pro- motion. 2017. Development of Practical Technique to Establish Fisheries Forensic Center; Research Project Report; KIMST: 2017. (In Korean)
-
26. Larsen T, Taylor DL, Leigh MB, O'Brien DM. 2009. Stable isotope fingerprinting: a novel method for identifying plant, fungal, or bacterial origins of amino acids. Ecology 90: 3526-3535.
-
27. McKinney CR, McCrea JM, Epstein S, Allen HA, Urey HC. 1950. Improvements in mass spectrometers for the measurement of small differences in isotope abundance ratios. Rev Sci Instrum 21: 724-730.
-
28. McMahon KW, McCarthy MD. 2016. Embracing variability in amino acid δ15N fractionation: mechanisms, implications, and appli- cations for trophic ecology. Ecosphere 7: e015
-
29. McMahon KW, Thorrold SR, Elsdon TS, McCarthy MD. 2015. Trophic discrimination of nitrogen stable isotopes in amino acids varies with diet quality in a marine fish. Limnol Oceanogr 60: 1076-1087.
-
30. Min J, Ha SY, Hur J, Shin K-H. 2019. Primary productivity and photosynthetic pigment production rates of periphyton and phytoplankton in Lake Paldang using 13C tracer. Korean J Ecol Env 52: 202-209.
-
31. Minagawa M, Wada E. 1984. Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48: 1135-1140.
-
32. Miura K, Goto AS. 2012. Stable nitrogen isotopic fractionation associated with transamination of glutamic acid to aspartic acid: implications for understanding 15N trophic enrichment in ecological food webs. Res Org Geochem 28: 13-17.
-
33. MOF, Ministry of Oceans and Fisheries. Ministry of Oceans and Fisheries 2020. Portal of Fisheries Information. Statistics on Marine Products Import and Export. Available online: http:// www.fips.go.kr/p/Main/ (accessed on 15 Oct. 2022).
-
34. Mzengereza K, Ishikawa M, Koshio S, Yokoyama S, Yukun Z, Shadrack RS, Seo S, Kotani T, Dossou S, Basuini MFE. 2021. Growth performance, growth related genes, digestibility, digestive enzyme activity, immune and stress responses of de novo camelina meal in diets of Red Seabream (Pagrus major). Animals 11: 3118.
-
35. Nam T-H, Ryu H-S, Kang T-W, Han Y-U, Kim J, Lee J, Hwang S, Kim K. 2019. Quantifying nitrogen source contribution ratios using stable isotopic method: Application of Bayesian mixing model. J Korean Soc Water Environ 35: 510-519.
-
36. O'Brien CL, Robinson SA, Pancost RD, Sinninghe Damsté JS, Schouten S, Lunt DJ, Alsenz H, Bornemann A, Bottini C, Brassell SC, Farnsworth A, Forster A, Huber BT, Inglis GN, Jenkyns HC, Linnert C, Littler K, Markwick P, McAnena A, Mutterlose J, Naafs BDA, Püttmann W, Sluijs A, van Helmond NAGM, Vellekoop J, Wagner T, Wrobel NE. 2017. Cretaceous sea-surface temperature evolution: Constraints from TEX86 and planktonic foraminiferal oxygen isotopes. Earth Sci Rev 172: 224-247.
-
37. Ohkouchi N, Chikaraishi Y, Close HG, Fry B, Larsen T, Madigan DJ, McCarthy MD, McMahon KW, Nagata T, Naito YI, Ogawa NO, Popp BN, Steffan S, Takano Y, Tayasu I, Wyatt ASJ, Yamaguchi YT, Yokoyama Y. 2017. Advances in the application of amino acid nitrogen isotopic analysis in ecological and biogeo- chemical studies. Org Geochem 113: 150-174.
-
38. Paulet Y-M, Lorrain A, Richard J, Pouvreau S. 2006. Experimental shift in diet δ13C: a potential tool for ecophysiological studies in marine bivalves. Org Geochem 37: 1359-1370.
-
39. Phillips DL, Inger R, Bearhop S, Jackson A, Moore J, Parnell A, Semmens B, Ward E. 2014. Best practices for use of stable isotope mixing models in food-web studies. Can J Zoo 92: 823-835.
-
40. Polito MJ, Trivelpiece WZ, Karnovsky NJ, Ng E, Patterson WP, Emslie SD. 2011. Integrating stomach content and stable isotope analyses to quantify the diets of Pygoscelid penguins. PLoS ONE 6: e26642.
-
41. Song B-Y, Gwak S, Jung M, Nam G, Kim NY. 2018. Tracing the source of methomyl using stable isotope analysis. Rapid Commun Mass Spectrom 32: 235-240.
-
42. Suh YJ, Shin K-H. 2013. Size-related and seasonal diet of the manila clam (Ruditapes philippinarum), as determined using dual stable isotopes. Estuar Coast Shelf Sci 135: 94-105.
-
43. Thomas F, Jamin E, Wietzerbin K, Guérin R, Lees M, Morvan E, Billault I, Derrien S, Rojas JMM, Serra F, Guillou C, Aursand M, McEvov L, Prael A, Robins RJ. 2008. Determination of origin of Atlantic salmon (Salmo salar): The use of multiprobe and multielement isotopic analyses in combination with fatty acid composition to assess wild or farmed origin. J Agri Food Chem 56: 989-997.
-
44. Trueman CN, MacKenzie KM, Palmer MR. 2012. Identifying migra- tions in marine fishes through stable-isotope analysis. Fish Biol 81: 826-847.
-
45. Velazco-Varga, J, Tomás-Vidal A, Hamdan M, Moyano López F, Jover Cerda M, Martínez-Llorens S. 2014. Influence of digest- ible protein levels on growth and feed utilization of juvenile meagre Argyrosomus regius. Aquacult Nutr 20: 520-531.
-
46. Wang YV, Wan AHL, Krogdahl Å, Johnson M, Larsen T. 2019. 13C values of glycolytic amino acids as indicators of carbohydrate utilization in carnivorous fish. Pee J 7: e7701.
-
47. Wang X, Li C, Wang M, Zheng P. 2018. Stable isotope signatures and nutritional sources of some dominant species from the PACManus hydrothermal area and the Desmos caldera. PLoS One 13: e0208887.
-
48. Whiteman JP, Smith EAE, Besser AC, Newsome SD. 2019. A guide to using compound-specific stable isotope analysis to study the fates of molecules in organisms and ecosystems. Diversity 11: 8.
-
49. Won E-J, Choi B, Lee CH, Hong S, Lee J-H, Shin K-H. 2020. Variability of trophic magnification factors as an effect of estimated trophic position: Application of compound-specific nitrogen isotope analysis of amino acids. Environ Int 135: 105361.
-
50. Won E-J, Kim SH, Go Y-S, Kumar KS, Kim M-S, Yoon S-H, Bayon G, Kim J-H, Shin K-H. 2021. Multi-elements isotope approach to assess the geographic provenance of Manila clams (Ruditapes philippinarum) via recombining appropriate elements. Foods 10: 646.
-
51. Won E-J, Yun H-Y, Lee D-H, Shin K-H. 2022. Application of compound-specific isotope analysis in environmental forensic and strategic management avenue for pesticide residues. Molecules 26: 10.3390.
-
52. Yokoyama H, Ishihi Y, ABO K, Takashi T. 2010. Quantification of waste feed and fish feces using stable carbon and nitrogen isotopes. Bull Fish Res Agen 31: 71-76.
-
53. Yun HY, Larsen T, Choi B, Won E-J, Shin K-H. 2022. Amino acid nitrogen and carbon isotope data: Potential and implications for ecological studies. Ecol Evol 12: e8929.
Copyright (C) 2024. The Korean Society of Marine Life Science