










JMLS 2020 December;5(2):58-63. http://jmls.or.kr Epub 2020 December 16
Copyright © 2020 by The Korean Society of Marine Life Science
Embryonic Development and Metamorphosis of the Ascidian Halocynthia aurantium
Gil Jung Kim; Department of Marine Molecular Bioscience, Gangneung-Wonju National University, Gangneung 25457, Korea
- Abstract
척삭동물문에 속하는 붉은멍게(Halocynthia aurantium)는 우렁쉥이와 같이 유용한 양식 품종으로 사료되지만, 발생과 생태 등 생물학적 특성에 대해 잘 알려지지 않았다. 본 연구에서는 붉은멍게 양식을 위한 기초자료를 얻기 위해 강원도 동해 연안에 서식하는 붉은멍게의 배발생을 조사하여 근연종인 우렁쉥이와 비교하였다. 그 결과, 붉은멍게의 수정부터 난할기, 낭배기, 신경배기의 배아 및 올챙이형 유생의 발달 단계 및 형태가 우렁쉥이와 매우 유사하였다. 붉은멍게의 수정란은 수온 11℃에서 부화까지 약 42.1시간이 소요되어 우렁쉥이의 40.9시간과 거의 유사하였다. 부화 후 어린개체로 변태하는 데 소요되는 시간도 두 종 사이에서 매우 유사하였다. 수온 11℃에서 부화한 두 종의 유생은 모두 약 23일이 경과해서 입수공과 출수공이 명확하게 구분되는 어린개체로 발생하였다. 수온 변화에 따른 발생 속도는 저온에서 느렸고 고온에서 빠른 결과를 나타냈다. 붉은멍게의 경우는 9℃에서 부화까지 평균 62.3시간, 11℃에서 42.1시간, 13℃에서 36.3시간이 소요되었다. 우렁쉥이의 경우는 평균 60.4시간, 40.9시간, 35.2시간이 소요되었다. 붉은멍게 배아의 대부분은 수온 15℃ 이상에서 정상적으로 발생이 이루어지지 않아 종묘생산 과정에 주의가 필요한 것으로 사료된다.
The ascidian Halocynthia aurantium (sea peach), which belongs to the phylum Chordata, is thought to be a valuable organism of aquaculture like H. roretzi (sea pineapple), but its biological characteristics such as development and ecology are not well known. In this study, in order to obtain basic data for H. aurantium farming, the development processes of H. aurantium inhabiting the east coast of Gangwon-do were investigated and compared with those of H. roretzi, a related species. As a result, the morphology and developmental stages of the fertilized eggs, embryos and larvae of H. aurantium were very similar to those of H. roretzi. Fertilized eggs of H. aurantium took about 42.1 hours to hatch at 11℃, almost similar to 40.9 hours of H. roretzi. The time required for larvae to metamorphose into juveniles after hatching was very similar between the two species. The hatched larvae of the two species became juveniles with oral and atrial siphons after 23 days at 11℃. Both types of embryos developed slowly in seawater at low temperatures and rapidly developed at high temperatures. Fertilized eggs of H. aurantium hatched in an average of 62.3 hours at 9℃, 42.1 hours at 11°C, and 36.3 hours at 13℃, whereas those of H. roretzi hatched in an average of 60.4 hours, 40.9 hours, and 35.2 hours. Most of H. aurantium embryos did not develop normally above 15℃, so it is thought that attention is needed in the seed production processes.
Keywords: Halocynthia aurantium(붉은멍게) Embryonic development(배발생) Meta- morphosis(변태) Water temperature(수온)
Correspondence to: Gil Jung Kim; Department of Marine Molecular Bioscience, Gangneung-Wonju National University, Gangneung 25457, Korea; E-mail : gjkim@gwnu.ac.kr
- Received
- 16 November 2020;
- Revised
- 18 November 2020;
- Accepted
- 3 December 2020.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
피낭류(tunicates)로도 불리는 멍게류(ascidians)는 미삭동물아문에 속하며 두삭동물아문 및 척추동물아문과 함께 척삭동물문을 구성한다. 올챙이 모양의 멍게류 유생은 척추동물의 원시조상과 유사한 형태인 것으로 추측된다(Katz, 1983; Satoh, 1994; Nishida, 2002). 멍게류는 발생과정에서 신경관을 유도하는 척삭 및 목과 머리 구조 발달에 중요한 인두열, 몸통에 발달한 근육 조직 등을 갖는다. 붉은멍게(Halocynthia aurantium)는 동해 북부, 일본 북부 태평양, 오호츠크해, 베링해, 태평양의 알래스카만 연안 등에 서식하는 것으로 보고되었으며, 강원도 삼척 연안에서 채집되었다는 보고가 있다(Van Name, 1945; Rho and Lee, 1991). 붉은멍게 유생의 형태 및 성체의 기본 구조는 우렁쉥이(H. roretzi)와 매우 유사하여 같은 속(Genus)으로 분류된다. 우렁쉥이(멍게)는 보통 수심 수 m 정도의 낮은 곳에서부터 30~40 m까지 분포하는 데 비하여 붉은멍게의 서식 수심은 10~100 m로 알려져 있으며 지역에 따라 편차가 있는 것으로 보인다. 강원도 연안에서는 수심 10~40 m 부근에 주로 서식하는 것으로 추정된다. 붉은멍게는 주로 원통형 모양으로 바다 속에서 높이 250 mm, 폭 90 mm를 넘는 대형 개체도 관찰되나, 채집한 개체를 공기 중에 놓아두면 평균 어른 주먹 크기만 하다. 붉은멍게의 체표면은 우렁쉥이와는 달리 돌기가 존재하지 않아 매끈하게 보이나 만지면 꺼끌꺼끌한 상태이다. 피낭의 색은 기본적으로 진한 붉은색이며 주황색과 검붉은색이 포함된다. 암반 등의 딱딱한 기질에 부착하여 서식하는 멍게류의 일반적인 특징과 같이 붉은멍게의 유생도 딱딱한 기질에 부착한 후 변태하여 성장하는 것으로 추정된다. 기질에 부착하는 조직의 형태는 우렁쉥이의 경우, 식물의 뿌리와 같은 구조가 넓고 촘촘하게 형성되지만, 붉은멍게는 몸의 밑 부분에 상대적으로 작은 크기로 자란다.
많은 해양동물처럼 멍게류의 산란은 일주기와 수온에 영향을 받는 것으로 알려져 있어 서식지에 따라 차이를 나타낸다. 강원도 연안에 서식하는 우렁쉥이의 산란기는 1월 전후, 동해 남부는 1월말에서 2월초, 남해안 통영 연안의 경우는 2월로 보고되었다(Bae, 1977; Kim et al., 2001). 하지만, 2017년 국립수산과학원이 발간한 멍게양식 매뉴얼에 따르면 기후 변화 등의 영향으로 남해안 우렁쉥이의 산란기가 12월말에서 1월까지로 한 달 이상 빨라졌다. 강원도 연안에 서식하는 붉은멍게의 산란기는 9~12월로 추정되지만, 아직 정확한 것은 추가적인 연구가 필요한 상황이다. 붉은멍게의 배발생과 변태과정에 대한 연구가 일부 수행되었지만(Lee et al., 2009), 자세한 내용은 추가적인 검토가 필요하다. 우렁쉥이는 우리나라의 주요 수산 양식 품종이지만, 기후 변화에 기인한 수온 상승과 환경변동으로 인해 잦은 폐사가 발생하여 양식어민들이 많은 어려움을 겪고 있다. 본 연구에서는 새로운 양식 멍게류의 개발을 위한 기초 연구로써 붉은멍게의 발생과 변태과정 및 수온의 영향에 관하여 조사하였다.
재료 및 방법
강원도 속초 연안에서 채집한 붉은멍게와 강릉 연안의 우렁쉥이를 연구에 사용하였다. 붉은멍게와 우렁쉥이는 수온 8℃, 24시간 형광등 조명 아래에서 사육하여 산란을 억제하였다. 각 개체를 별개의 투명용기에 담아 수온 11℃에서 7시간 이상 차광한 후, 다시 빛을 공급하는 방법으로 산란과 방정을 유도하였다. 난자에 다른 개체에서 유래한 정자 희석액을 섞어줌으로써 수정란을 얻었다. 붉은멍게의 경우, 우렁쉥이와 유사한 방법에 의해 산란과 방정이 제대로 유도되지 않았다. 간출자극도 붉은멍게의 산란과 방정 유도에 유의미한 효과를 나타내지 않았다. 실험과정에서 우렁쉥이와 같은 산란 유도 방법에 의해 일부 방출된 난자와 생식소 해부를 통해 얻은 정자를 섞어 붉은멍게의 수정란을 얻었다. 붉은멍게와 우렁쉥이의 난자와 정자를 각각 섞은 다음 약 1시간 후에 수정막이 부풀어 오르는 것을 확인한 후, 직경 100 μm의 거름망을 사용하여 해수로 여분의 정자를 씻어냈다. 수정란은 저온배양기에서 9℃, 11℃ 및 13℃에서 사육하였다. 총 3회 실시한 실험의 결과를 평균하여 발생 시간을 결정하였으며, 각 실험의 모집단(n)은 20개 이상이었다. 부화한 유생을 거름막으로 걸러 바닷물에 남아 있는 난황막을 제거한 후, 직경 60 mm의 petri dish에 30마리를 담아 11℃에서 변태과정을 조사하였다. 13℃ 이상의 수온에서 붉은멍게의 배아가 정상적으로 발생하는지 조사하기 위하여 수정란을 수온 15℃ 및 20℃에서 사육하였다. 배아와 유생의 발생은 실체 현미경(OLYMPUS SZX 12)을 사용하여 관찰하였고, 디지털 카메라(OLYMPUS microscope digital camera DP25)를 이용하여 발생과 변태의 각 단계를 촬영하였다.
결 과
1. 붉은멍게의 배발생과 부화
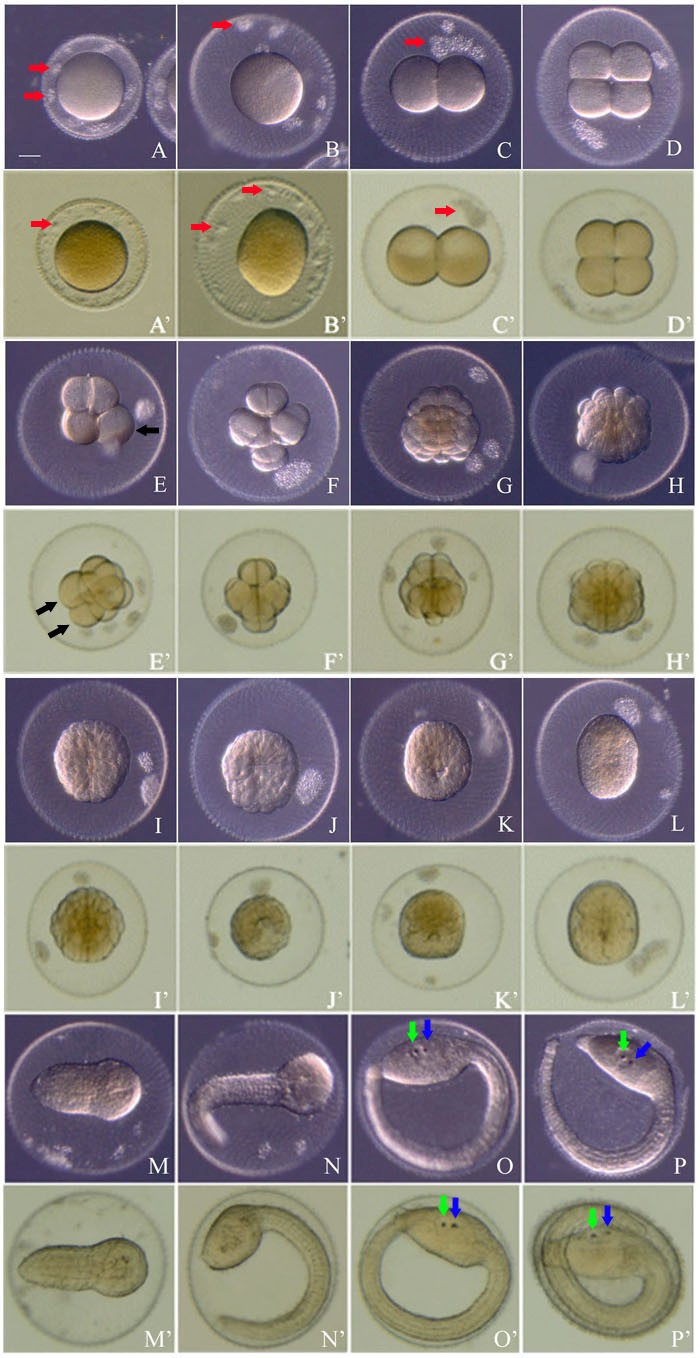
붉은멍게의 수정란을 수온 11℃에서 배양하며 발생 단계 별 소요 시간을 조사하였다(Fig. 1; Table 1). 붉은멍게의 알은 우렁쉥이 알에 비하여 노란색이 현저하게 적어 약하게 노란빛이 감도는 연한 회색으로 보였다. 붉은멍게 난자의 난황막(vitelline membrane) 외부에는 난포세포가 단층으로 난을 감싸고 있다(Fig. 1A). 정자가 난자에 결합한 후, 약 10분만에 난황막이 부풀어 오르기 시작하여 위란강이 확장되었다(Fig. 1B). 수정 후 약 2.1시간에 수정란은 동-식물극 축을 따라 난자의 세포질을 반으로 나누는 첫 번째 세포질분열을 시작하였다. 세포질분열이 시작되어 10분 정도 경과한 수정 후 2.2시간에 2세포기 배아가 되었다(Fig. 1C). 이후 각 세포기의 세포질분열 시간은 거의 유사하여 세포질분열이 시작되어 완료되기까지 약 10분이 소요되었다. 수정 후 약 3.2시간에 4세포기가 되었고, 이때 두 번째 난할면은 첫 난할면과 직각이며 동-식물극 축을 따라 일어난다(Fig. 1D). 이어서 동-식물극 축에 직교하여 세 번째 난할이 일어나 수정란은 약 4.3시간에 8 세포기 배아가 되었다(Fig. 1E). 수정 후 약 5.3시간이 경과하면 네 번째 난할이 일어나 배아는 16세포기가 되었다(Fig. 1F). 이어 난할이 계속되어 32세포기(Fig. 1G)와 64세포기(Fig. 1H)를 거쳐 수정 후 약 11시간에 낭배형성과정이 시작되는 110 세포기가 되었다(Fig. 1I).
낭배형성을 마친 배아는 신경배아기를 거쳐 수정 후 약 24시간에 초기 미충형 유생기(early tailbud stage)가 되었다(Fig. 1M). 중기 미충형 유생기 이후부터 꼬리가 더욱 신장함에 따라 꼬리는 난황막 안에서 휘어지고, 약 30시간 발생한 후기 미충형 유생의 머리에서는 안점(ocellus)과 평형기(otolith) 색소세포가 처음으로 관찰되었다(Fig. 1O). 수정 후 약 42.1시간에 유생의 움직임이 격렬해지면서 난황막을 뚫고 부화하였다(Fig. 1P). 부화 직후에는 꼬리가 휘어져 있어 움직임이 활발하지 못하지만 30 여분 이내에 꼬리가 펴지면서 올챙이형 유생은 활발하게 유영을 시작하였다.
붉은멍게와 우렁쉥이 간에 난할의 방식과 초기 발생 소요 시간은 매우 유사한 결과를 나타내었다(Fig. 1; Table 1). 수온 11℃에서 붉은멍게 배아는 우렁쉥이 배아에 비해 약간 발생이 느렸지만 거의 유사한 발생 속도와 매우 비슷한 배아 형태를 나타냈다. 부화 직후 붉은멍게 올챙이형 유생의 크기는 머리에서 꼬리까지 평균 2.1±0.1 mm이고, 머리의 크기는 0.22±0.02 mm이다. 반면 우렁쉥이는 길이 2.4±0.1 mm, 머리는 0.26±0.02 mm로 붉은멍게의 유생이 우렁쉥이보다 길이 약 0.3 mm, 머리는 약 0.4 mm 정도 작은 것으로 측정되었다.
2. 붉은멍게 유생의 변태
수온 11℃에서 부화한 붉은멍게 유생의 변태과정을 조사하였다(Fig. 2; Table 2). Halocynthia 속의 멍게는 유생의 밀도에 따라 변태 속도가 영향을 받는 것으로 알려져 있다(Hirano and Nishida, 2000). 따라서 실험 방법에서 언급한 것처럼 부화한 유생을 직경 60 mm의 petri dish에 30마리씩 담아 변태과정을 조사하였다. 부화 후 4일째에 꼬리가 시작되는 부분에서 척삭과 근육세포 등 꼬리를 구성하는 내부 조직이 몸통으로 흡수되기 시작한다(Fig. 2A, B). 부화 후 약 6일이 경과하면 꼬리는 몸통으로 완전히 흡수되고 꼬리 부분의 피낭 흔적이 관찰되며, 돌출된 부착돌기를 명확하게 확인할 수 있다(Fig. 2C, D). 부착돌기는 계속적으로 성장하여 부화 후 약 2주일 경에 변태 중인 유생의 몸과 거의 같은 길이로 자란다(Fig. 2E). Fig. 2의 변태하는 유생들은 부착돌기를 자세하게 나타내기 위해 용기의 바닥에서 떼어낸 모습이며, 자라기 시작한 유생의 부착돌기는 바닥에 단단히 붙게 된다. 부화 후 약 3주가 경과한 유생은 변태를 완료하고 입수공과 출수공을 갖는 어린개체(juvenile)가 된다(Fig. 2F). 부화한 유생이 변태하는 속도도 붉은멍게와 우렁쉥이 사이에서 유사했으며 형태적으로도 매우 닮았다(Fig. 2; Table 2).
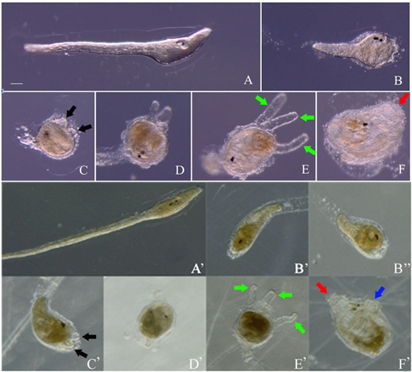
Fig. 2. Metamorphosis of Halocynthia aurantium (A-F) and Halocynthia roretzi (A'-F'). (A, A') Tadpole larvae that begin to metamorphose. (B, B', B") Premeta- morphic larvae with a short tail. (C, C') Premetamorphic larvae 6 (C) and 8 (C') days old after hatching. Black arrows in- dicate adhesive papillae. (D, D') Premeta- morphic larvae 13 (D) and 12 (D') days old after hatching. (E, E') Premetamorphic larvae 16 days old after hatching. Green arrows indicate protruding adhesive pa- pillae to the maximum. (F, F') Juveniles with oral (incurrent) siphon (red arrows) and atrial (outcurrent) siphon (blue arrow). Scale bar, 100 μm.

3. 수온에 따른 붉은멍게와 우렁쉥이의 부화 시간 비교
멍게류는 수온에 따라 발생 속도가 크게 영향을 받는다. 수온 9℃, 11℃ 및 13℃에서 붉은멍게와 우렁쉥이 수정란의 부화 소요 시간을 조사하였다(Table 3). 수온 9℃에서 붉은멍게 수정란은 62.3시간에 부화하였고, 우렁쉥이 수정란도 유사한 60.4시간에 부화하였다. 수온 11℃에서 붉은멍게와 우렁쉥이 수정란은 각각 42.1시간과 40.9시간에 부화하였고, 수온 13℃에서는 각각 36.3시간과 35.2시간에 부화하였다. 예상한 것처럼 수온이 낮을수록 발생이 느리게 진행되었다. 붉은멍게의 수정란을 수온 15℃에서 발생시키면 비정상적인 난할, 신경관 닫힘 이상 또는 꼬리 휨 등의 형태 이상을 나타내는 비율이 크게 높아 졌다. 수온 13℃에서 수정란의 80% 이상이 정상적인 유생으로 발생했지만, 수온 15℃에서는 정상 발생의 비율이 30% 이하로 관찰되었다. 또한, 우렁쉥이와 붉은멍게의 수정란은 수온 20℃에서 거의 대부분 정상 발생하지 않았다.

고 찰
자웅동체인 멍게류의 많은 종들은 한 개체에서 유래하는 정자와 난자 사이에는 수정이 일어나지 않는다(Fuke, 1983; Satoh, 1994; Jiang and Smith, 2005). 우렁쉥이와 마찬가지로 붉은멍게에서도 출수공을 통하여 산란된 정자와 난자는 다른 개체에서 유래된 상대 배우자와 수정되었다. 붉은멍게에서 정자가 난자에 침투한 다음 약 10분 이내에 난황막이 부풀어 오르기 시작하여 첫 세포분열 시작 시점까지 위란강이 확장되었다(Fig. 1B). 이 공간은 배아의 정상적인 발생을 위하여 필요한 것으로 공간의 확장이 부족하면 꼬리가 굽어지는 비정상적인 유생이 출현하는 비율이 크게 증가한다. 이 난황막의 확장에는 난황막에 존재하는 난포세포가 중요한 기능을 하는데 실험적으로 난포세포를 제거한 난자는 난황막의 확장을 나타내지 않는다. 따라서 종묘생산과정에서 비정상적인 유생의 비율을 낮추기 위해 위란강이 확장되는 수정 직후 2시간 가량 수정란에 충격이 가해지지 않도록 주의가 필요하다. 우렁쉥이에서도 난황막의 확장은 발생에 있어 중요하며 붉은멍게와 유사한 특징을 나타낸다(Satoh, 1994). 그러나 유령멍게류(Ciona 속)의 수정에서는 뚜렷한 난황막의 확장이 관찰되지 않아 서로 다른 모습을 보인다(Yoshida et al., 1993; Jiang and Smith, 2005).
전통적인 난할의 구분 방법에 따르면 붉은멍게의 첫 난할과 두 번째 난할은 경할이고 세 번째 난할은 위할에 해당한다. Fig. 1E, E'와 같이 붉은멍게와 우렁쉥이 8 세포배아의 모습은 성게 8 세포배아의 모습과 달리 외관상으로 동-식물극과 앞-뒤 및 좌-우 축의 방향을 구분할 수 있는 독특한 모습이다. 이러한 형태는 배아의 뒤쪽 식물극 할구(B4.1)에 위치하는 CAB (centrosome attracting body)라는 세포의 크기와 분열 방향을 결정하는 세포 내 기구의 특특한 배열과 작용 때문이다(Nishikata et al., 1999; Negishi and Nishida, 2017). 이처럼 붉은멍게 수정란의 크기, 배아의 형태, 난할 방식은 우렁쉥이와 거의 유사하였다. 난자의 세포질은 붉은멍게에 비하여 우렁쉥이가 노란색이 진한 경우가 많았지만, 개체에 따라 우렁쉥이 난자도 붉은멍게처럼 연한 회색인 경우가 존재했다.
이(Lee et al., 2009) 등은 붉은멍게의 수정에서부터 부화까지 소요 시간이 12℃에서 19.5시간이라고 보고하였다. 또한, 입수공과 출수공의 형성이 만 4일 이내에 시작된다고 하였다. 이것은 수온 1℃의 차이를 고려하더라도 본 실험의 결과와 매우 큰 차이를 나타낸다. 또한, 그들은 수온 20℃ 이상에서 붉은멍게의 발생을 조사하였는데 본 실험 결과에 따르면 우렁쉥이와 붉은멍게의 수정란은 수온 20℃ 이상에서 정상적인 발생을 거의 하지 않는다. 붉은멍게의 수정란은 수온 15℃에서 발생 장애가 현저하게 높은 비율로 나타났다. 이에 대한 검증은 앞으로 다른 연구그룹에서 확인할 필요가 있다. 유령멍게(Ciona intestinalis)의 경우는 수온 18℃에서 수정 후 약 1시간 경에 첫 세포분열을 하고, 수정 후 약 17.5시간에 올챙이형 유생으로 부화한다. 부화한 유생은 평균 3시간 후 기질에 부착하여 부화 후 12시간 경에 꼬리의 흡수가 완료되는 것으로 보고되었다(Matsunobu and Sasakura, 2015). 유령멍게의 꼬리 흡수에 걸리는 시간은 붉은멍게와 우렁쉥이에 비해 2배 이상 빠른 속도이다.
우렁쉥이는 국내 주요한 양식 품종이지만 물렁증의 발생 및 여름철 고수온과 빈산소의 영향으로 생산량 변동이 심하여 양식어민들에게 많은 어려움을 준다. 동해 연안에 서식하는 붉은멍게의 양식이 안정적으로 이루어지면 멍게 양식어민들에게 큰 경제적 이익이 될 것으로 판단한다. 본 실험의 결과가 붉은멍게 양식을 위한 종묘생산 과정에서 유용한 기초자료가 될 것으로 사료한다.
- References
-
1. Bae SH. 1977. The spawning Halocynthia roretzi (Drasche) and the influences from water temperature. Kunsan Fisheries College, Research Reports 11: 65-69.
-
2. Fuke MT. 1983. Self and non-self recognition between gametes of the ascidian, Halocynthia roretzi. Wilehm Roux Arch Dev Biol 192: 347-352.


-
3. Hirano T, Nishida H. 2000. Developmental fates of larval tissues after metamorphosis in the ascidian, Halocynthia roretzi. II. Origin of endodermal tissues of the juvenile. Dev Genes Evol 210: 55-63.
-
4. Jiang D, Smith WC. 2005. Self- and cross-fertilization in the solitary ascidian Ciona savignyi. Biol Bull 209: 107-112.
-
5. Katz MJ. 1983. Comparative anatomy of the tunicate tadpole, Ciona intestinalis. Biol Bull 164: 1-27.
-
6. Kim BS, Bang JD, Ryu HY, Hong JP, Chung EY. 2001. Gametogenesis, Gonadal development and maturation of the sea squirt, Halocynthia roretzi. Dev Reprod 5: 137-144.
-
7. Lee C, Park MW, Lee CS, Kim SK, Kim WK. 2009. Effects of Temperature and Salinity on Development of sea peach Halocynthia aurantium. J Environ Sci 18: 1171-1179.
-
8. Matsunobu S, Sasakura Y. 2015. Time course for tail regression during metamorphosis of the ascidian Ciona intestinalis. Dev Biol 405: 71-81.
-
9. Negishi T, Nishida H. 2017. Asymmetric and Unequal Cell Divisions in Ascidian Embryos. Results Probl Cell Differ 61: 261-284.
-
10. Nishida H. 2002. Specification of developmental fates in ascidian embryos: molecular approach to maternal determinants and signaling molecules. Int Rev Cytol 217: 227-276.
-
11. Nishikata T, Hibino T, Nishida H. 1999. The centrosome-attracting body, microtubule system, and posterior egg cytoplasm are involved in positioning of cleavage planes in the ascidian embryo. Dev Biol 209: 72-85.
-
-
13. Satoh N. 1994. Developmental biology of ascidians. Cambridge University Press, New York, pp 3-149.
-
14. Van Name WG. 1945. The North and South American ascidians. Bull Amer Mus Nat Hist 84: 1-476. (p. 362)
-
15. Yoshida M, Inaba K, Morisawa M. 1993. Sperm chemotaxis during the process of fertilization in the ascidians Ciona savignyi and Ciona intestinalis. Dev Biol 157: 497-506.
Copyright (C) 2024. The Korean Society of Marine Life Science