










JMLS 2020 June;5(1):17-24. http://jmls.or.kr Epub 2020 June 16
Copyright © 2020 by The Korean Society of Marine Life Science
Characterization of Heat Shock Protein 70 in Freshwater Snail, Semisulcospira coreana in Response to Temperature and Salinity
Seung Rae Park; Department of Marine Biotechnology, Gangneung-Wonju National University, Gangneung 25457, Korea
Young Kwang Choi; Department of Marine Biotechnology, Gangneung-Wonju National University, Gangneung 25457, Korea
Hwa Jin Lee; Department of Marine Biotechnology, Gangneung-Wonju National University, Gangneung 25457, Korea
Sang Yoon Lee; The East Coast Research Institute of Life Science, Gangneung-Wonju National University, Gangneung 25457, Korea
Yi Kyung Kim; Department of Marine Biotechnology, Gangneung-Wonju National University, Gangneung 25457, Korea;The East Coast Research Institute of Life Science, Gangneung-Wonju National University, Gangneung 25457, Korea
- Abstract
참다슬기 아가미 조직으로부터 heat shock protein 70 유전자를 분리 · 동정하였다. 참다슬기HSP70 cDNA의 open reading frame (ORF)는 1,917 bp로 639개의 아미노산을 암호화하여 분자량은 약 70 kDa으로 예측되었다. 생물정보학 배열분석에 의해 HSP 유전자 기능과 관여되어 있는 3가지 주요 signature motifs와 보존된 도메인을 확인하였다. 계통학적 분석을 통하여 참다슬기 HSP70 유전자는 왕우렁이 Pomacea canaliculate와 같은 클러스트에 포함된다는 사실을 확인하였다. 수온 및 염분 변화에 따라, 참다슬기 HSP70 mRNA 유전자 레벨은 유의적으로 증가하였으며(p < 0.05), 이는 외부자극요인을 파악할 있는 분자생물학적 마커로서 활용될 수 있을 것으로 사료된다.
We have identified a heat shock protein 70 gene from freshwater snail, Semisulcospira coreana. The freshwater snail HSP70 gene encode a polypeptide of 639 amino acids. Based on bioinformatic sequence characterization, HSP70 gene possessed three classical signature motifs and other conserved residues essential for their functionality. The phylogenetic analysis showed that S. coreana HSP70 had closet relationship with that of golden apple snails, Pomacea canaliculata. The HSP70 mRNA level was significantly up-regulated in response to thermal and salinity challenges. These results are in agreement with the results of other species, indicating that S. coreana HSP70 used be a potential molecular marker in response to external stressors and the regulatory process related to the HSP70 transcriptional response can be highly conserved among species.
Keywords: Semisulcospira coreana(참다슬기) Heat shock protein 70(열충격단백질) Temperature(수온) Salinity(염분)
Correspondence to: Yi Kyung Kim; Department of Marine Biotechnology, Gangneung-Wonju National University, Gangneung 25457, Korea; E-mail : yikyung1118@gwnu.ac.kr;
- Received
- 13 April 2020;
- Revised
- 16 April 2020;
- Accepted
- 27 April 2020.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
담수산 참다슬기, Semisulcospira coreana는 복족강(gastropoda)에 속하며, 한국, 일본, 중국 남부 그리고 타이완 등을 포함한 동아시아 대부분의 지역에서 널리 서식하고 있는 종이다(Lee et al., 2007; Köhler, 2017). 우리나라에서는 건강보조식품으로서 각광받으며 국내소비량이 점차 증가하고 있으며, 특히 국내산 선호도가 높아 다슬기는 경제성이 높은 고급기호품종으로 양식산업에서 경쟁력이 있는 대상종으로서 인지되고 있다(Song and Kang, 2016). 그러나 현재 수질오염과 무분별한 개발 등으로 인한 담수생태계 감소 및 무분별한 남획 등으로 인하여 다슬기 자원량이 급격하게 줄어들고 있다. 이러한 배경으로 인하여 다슬기 양식 기술개발 필요성이 대두되어, 인공종묘생산, 사료 기술개발 등 다양한 연구가 시도되고 있다.
수생동물의 아가미 조직은 외부환경과 접하면서 체표면이 가장 넓은 조직이며, 미생물학적 감염 혹은 조직 손상에 대한 1차 방어를 담당하는 점막표층으로 둘러싸여 있다(Tort et al., 2003). 이외에도 아가미 조직은 가스교환, 삼투조절, 질소 노폐물 배출 및 산-염기 균형 등을 포함하는 생리작용에 중요한 역할을 하는 것으로 알려져 있다(Evans et al., 2005). 세포는 주위환경으로부터 자극을 받으면 세포의 본래 기능을 유지하고 어려워지고, 몇몇 단백질에 의해 세포의 원래 기능이 유지되도록 발현된다. 열충격단백질(Heat shock proteins, HSPs)은 다양한 생물학적 혹은 비생물학적 요인들에 반응하며 발현하는 단백질로서 스트레스 단백질로도 알려져 있다(Srivastava, 2002). 스트레스가 가해지지 않은 경우에도 세포 내 단백질 접힘, 기능 저하, 단백질 구조 변형을 방지하는 등의 분자 샤페론으로서 작용하고 있으며, 세포의 항상성 유지를 돕는 역할을 한다(Hightower, 1991; Kim et al., 2013). HSP 단백질은 분자량에 따라 HSP90 (85~90 kDa), HSP70 (68~73 kDa), HSP60, HSP47 그리고 저분자량 HSPs (16~24 kDa)로 분류된다(Feder and Hofmann, 1999; Trent, 1996). 그 중에서도 HSP70 family는 가장 잘 보존되어 있으며, 세균에서 식물, 그리고 포유류에 이르기까지 다양한 생물체 내에서 많은 화학적 혹은 생물학적 자극을 통해서 발현되는 것으로 알려져 있다(Song et al., 2014). HSP70 family 중에서 가장 많이 연구된 유전자는 Hsc73 (heat shock cognate 73 kDa protein)과 HSP70이다. Hsc73 유전자는 대부분의 세포에서 발현되며, 발현 유도가 매우 낮은 특징을 가지며, 스트레스에 노출되었을 경우에는 핵과 인으로 이동한다(Welch and Feramisco, 1984). 이에 반해 HSP70은 정상적인 상태에서는 발현이 적다가 자극을 받았을 때만 급격하고 반응하여 상당양의 증가를 나타낸다. 세포질 HSP70은 부분적으로 합성된 펩타이드 서열에 결합하므로써 기능을 차단시키거나 단백질이 합성되면 HSP70은 떨어져 나와 3차구조로 접히게 한다(Morimoto, 1998). 이외에도 직접적으로 세포사멸을 억제하는 기능을 가지는 것으로 알려졌다. 이와 같이 자극이 가해지면 세포 내에서 외부자극에 대한 변화를 탐지해 낼 수 있는 생체지표로서도 많이 활용되고 있으며, 세포 내 기관에 존재하고 있어 스트레스에 의한 손상으로부터 세포를 보호하는 기능을 수행하고 있다. 복족류 HSP 유전자의 분자적 특징과 발현에 대한 정보는 유럽전복 Haliotis tuberculate, 참전복 H. discus hannai, 달팽이 Biomphalaria glabrata, 애기삿갓조개 Cellana toreuma 같은 종에 국한되어 있다(Laursen et al., 1997; Cheng et al., 2007; Farcy et al., 2007; Han et al., 2013). 다슬기 양식 현장에서는 다슬기 어미의 이동이나 자연에서 유입되는 거머리(Glossiphonia sp.)를 제거하는데 많은 어려움을 겪고 있다. 거머리가 한번 발생하면 구제하기가 어렵고, 다슬기와 거머리가 모든 약품이나 환경내성이 비슷하여 거머리만 선택적으로 폐사시키기 쉽지 않다(Kim et al., 2009). 거머리를 제거할 수 있는 방안을 강구하기 위하여 거머리가 염분도 12.96 psu까지 생존 및 적응할 수 있다는 기존 연구 결과(Mcattleadanmohanrao, 1960)를 근거로 하여 담수산 참다슬기의 염분 적응력을 파악하고자 하였다. 다양한 환경 스트레스를 제어하는데 중요한 역할을 하는 다슬기 HSP 단백질에 대한 분자생물학적 구명 및 환경 변화에 대한 참다슬기의 생리학적 해석에 대한 연구는 미비한 실정이다. 따라서, 본 연구에서는 참다슬기 아가미 세포에서 발현되는 HSP70 유전자의 분자생물학적 특성을 구명하고 수온 및 염분 변화에 따른 HSP70 유전자 발현 양상을 분석하였다. 이러한 결과들은 참다슬기의 환경에 따른 생리학적 지표를 설정하는 기초자료로 이용될 수 있을 것으로 사료된다.
재료 및 방법
1. 실험동물 및 실험조건
본 연구에 사용된 개인양식장에서 구입한 참다슬기를 강릉원주대학교 실험실로 운반하여 30마리 정도 순환수조에서 넣어 일주일 동안 순치시켰다. 수온 변화의 조건을 설정하기 위해서 물환경정보시스템(http://water.nier.go.kr/main/mainContent.do)에서 2018년 금강 월평균 수온을 조사하였다. 참다슬기의 산란이 시작되는 6월의 월평균 수온인 24℃를 대조구로, 월평균 수온이 가장 높은 8월의 평균 수온인 30℃를 고수온으로 정의하였다. 수온 변화는 1시간당 2℃씩 상승하도록 설정하여, 24℃(대조구), 고수온 조건(30℃ 및 30℃ 도달한 후 24시간)으로 나누어 샘플링을 실시하였으며, 각 3마리 다슬기의 아가미 조직을 채취하여 -80℃ 냉장고에 보관하였다. 다슬기의 염분 적응력을 시험하기 위해서 거머리의 염분 적응력(Mcattleadanmohanrao, 1960)을 근거로 하여 담수조건(0 psu)을 대조구로 하여, 다슬기 개체를 5개의 실험구 4, 6, 8, 10 psu로 나누어 24시간 동안 노출시켰다.
2. 참다슬기 HSP70 유전자 클로닝과 분자적 특징분석
실험실 내 참다슬기 NGS database에서 기존에 알려진 HSP과 높은 상동성을 나타내는 NGS clones을 수집하여 확보된 HSP70 cDNA 전체 배열을 BLAST를 이용하여 확인하였다. Specific primer pair set (FW: 5'-ATGGGATCGAATAAAGCTCC-3', RV: 5'-TTAGTCGA- CCTCCTCCACGG-3')에 의한 RT-PCR 증폭을 통하여 HSP70 cDNA 전체 배열을 확인하였다. 다슬기와 다른 종과의 상동성을 비교분석하기 위해서 BLAST searches를 검색하였다. CLUSTAL W 프로그램을 이용하여 Multiple sequence alignments를 수행하였다(Thompson et al., 1994). HSP70의 단백질 도메인은 InterPro software을 이용하여 분석하였다(Hunter et al., 2009). Molecular Evolutionary Genetics Analysis (MEGA) program (ver. 5.2)을 이용하여 neighbor-joining 방법으로 계통수 분석을 실시하였다.
3. Total RNA 추출 및 cDNA 합성
자극에 따른 참다슬기 HSP70 유전자 mRNA 발현를 분석하기 위해 대조구 및 실험구 다슬기 개체에서 아가미 조직을 분리하여 RNAiso Reagent (Takara Bio, Shiga, Japan)을 이용하여 total RNA를 추출하였다. Nanodrop ND-2000 (Thermo Scientific, Waltham, USA) 분광광도계를 사용하여 흡광도를 측정한 후 RNA를 정량하였다. RNeasy Plus Mini Kit (Qiagen, Hilden, Germany)와 RNase-free DNase set (Qiagen)을 이용하여 추가적인 정제작업을 진행하였다. cDNA 합성은 PrimeScript RT reagent kit (Takara)을 사용하여 제조자의 매뉴얼에 따라 RNA 역전사 반응을 유도하였다.
4. Quantitative real-time RT-PCR (qPCR)
참다슬기의 아가미 조직 내 HSP70 유전자의 발현분석을 위해서 Thermal Cycler DiceTM real-time PCR system (Takara)과 SYBR premix ExTaqII Kit (Takara)을 사용하였다. HSP70 증폭에는 oligonucleotide primer pairs, qscHSP70-FW (5'-AGAGTTTGAGGACAAG- CTGC-3')와 qSCHSP70-RV (5'-TGATCTAATGTCCAATGCGC-3')을 이용하였고, 증폭되는 사이즈는 188 bp이다. Reference gene으로서 참다슬기 ribosomal protein L7 (RPL7) (FW: 5'-CATTGCTCACGATGTCG- ATC-3', RV: 5'-TGTCTTCACTGTTGACGGTG-3')을 사용하였으며, normalization control로서 각 실험구간 상대정량을 실시하였다. Real-time PCR 조건은 초기 변성으로 95℃ 30초간 1 cycle, 이어서 95℃에서 5초, 60℃에서 30초를 45 cycles로 수행하였다. 유전자의 실험 결과는 Schmittgen and Livak (2008) 방법에 따라 2-ΔΔCt 값으로 계산하였다.
5. 통계분석
실험 결과는 mean ± standard error (SEM)로 나타냈으며 SPSS 25.0 software (SAS Inc., Cary, NC, USA)을 이용하여 one-way ANOVA 및 Duncan's multiple range tests를 실시하여 유의성(p < 0.05)을 분석하였다.
결과 및 고찰
1. 참다슬기 heat shock protein (HSP) 70 cDNA 유전자의 분자유전학적 특성
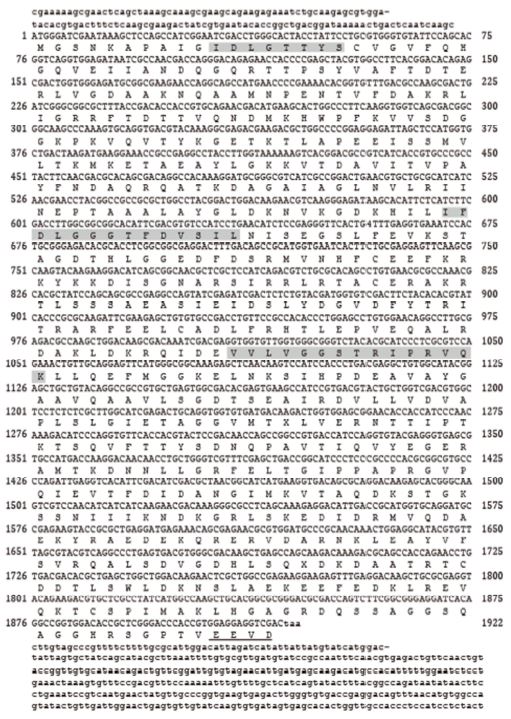
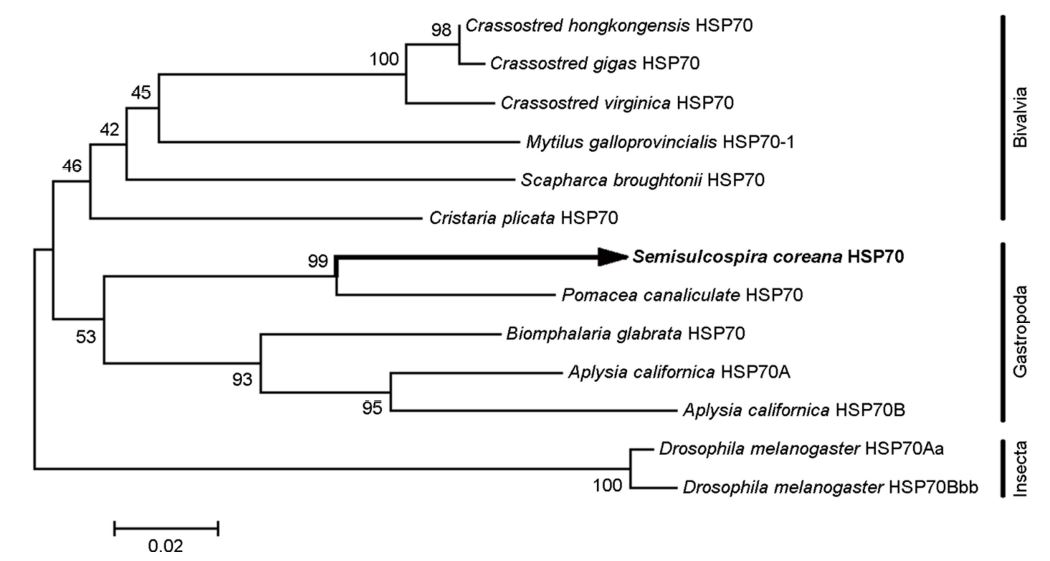
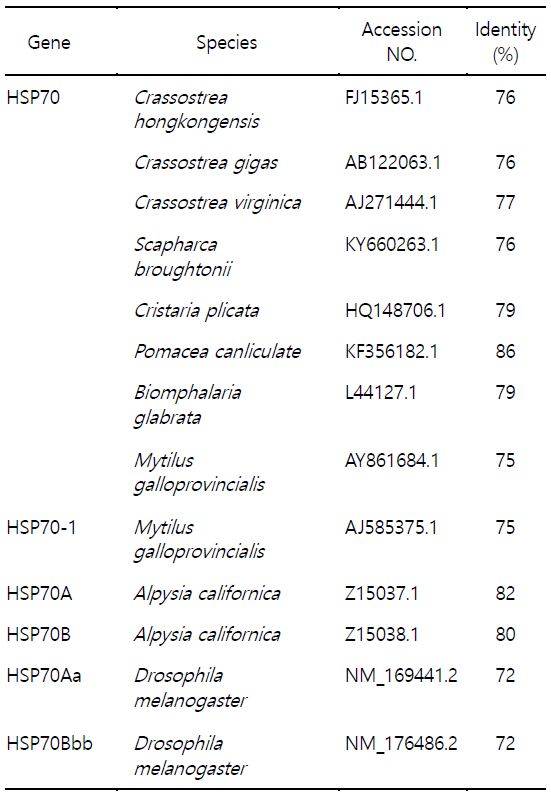
참다슬기의 아가미 조직으로부터 heat shock protein 70 (scHSP70) cDNA를 분리하여 분자생물학적 특징을 분석하였다. HSP70 cDNA의 open reading frame (ORF)는 1,917 bp로 639개의 아미노산을 암호화하여 분자량은 약 70 kDa으로 예측되었다(Fig. 1). 참다슬기 HSP70의 deduced amino acid sequences 정보에서 3개의 signature motifs (11-18 aa, IDLGTTYS; 199-212 aa, IFDLGGGTFDVSIL; 337-351 aa, VVLVGGSTRIPRVQK)가 확인되었으며 이는 모든 HSP70 family에서 매우 잘 보존된 영역으로 알려져 있다. Cytoplasmic HSP70의 C 말단에는 consensus EEVD tail (636-639 aa)이 확인되었다. 이외에도 추가적으로 putative adenosine triphosphate (ATP)-guanosine triphosphate (GTP) binding domain (AEAYLG)와 potential nonorganelle eukaryotic consensus motif (RARFEEL)와 gylocosylation domain (NKSI)이 확인되었다. Multiple sequence alignment 결과, 왕우렁이 P. canaliculate와 민물달팽이 Biomphalaria glabrata와는 84.7%와 79%, 높은 상동성을 나타내었고 다른 종과도 유사한 상동성을 나타내었다(72~80%) (Table 1, Fig. 2). 뿐만 아니라 배열분석과 구조적 특징을 통해서 참다슬기 아가미 조직으로부터 새롭게 동정된 HSP은 전형적인 열유도성 HSP70 family에 속하는 유전자로 확인되었다. 그러나, HSP70의 구조적인 특성 중 하나이며, 단백질 co-chaperon 상호작용을 조절하는데 관여한다고 알려진 GGMP tetrapeptide repeats의 존재는 확인되지 않았다(Piano et al., 2005). 계통수 분석에서도 다른 복족류 HSP70과 같은 cluster 안에 포함되는 것을 확인하였다(Fig. 3).

2. 수온 및 염분변화에 따른 참다슬기 HSP70 발현 분석
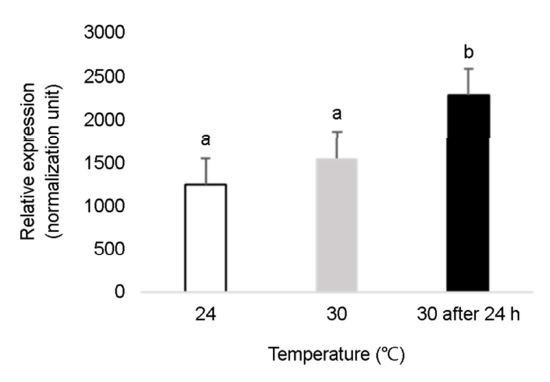
환경스트레스로 인한 참다슬기의 생리적 변화를 검토하기 위해 수온 및 염분 변화에 따른 HSP70 유전자 변화를 검토하였다(Figs. 4, 5). 참다슬기 HSP70 발현양은 대조군(24℃) 및 30℃에서는 유의적인 차이를 나타내지 않았으나, 30℃에 도달한 후 24시간 후에는 약 2배 이상 유의적인 높은 발현양을 나타내었다(Fig. 4). HSP70 mRNA은 열충격을 가하지 않은 상태, 적정수온 범위에 있는 다슬기 아가미 조직에서도 열충격을 가한 30℃ 구간에서 유사한 발현양이 확인되었다. 이는 HSP70 유전자가 정상적인 생리학적 조건에서도 분자 샤페론으로 작용하여 단백질의 합성, 접힘 등을 돕는 등 이는 생물체 내 생리작용 혹은 세포 항상성을 유지하는데 에너지를 소모하고 있다는 사실을 예측할 수 있다. 다슬기의 최적 수온, 24℃ 구간과 고수온 30℃에 도달한 시점에서는 HSP70 유전자의 유의적인 차이를 나타내지 않았으나, 고수온에 도달한 후 24시간이 경과된 시점에서 유의적인 발현증가를 나타내었다. 이는 참다슬기의 고온 스트레스에 대한 효과를 경감시키기 위한 반응으로 파악되며, 이는 생체 내에서 HSP 발현양과 열적응력(heat tolerance) 사이에서 양의 상관관계가 있다는 것을 시사하고 있다. 이러한 증가하는 경향은 유럽전복(Farcy et al., 2007), 지중해담치 Mytilus galloprovincialis (Cellura et al., 2006), 넓적굴 Ostrea edulis (Piano et al., 2002), 둥근전복 Haliotis discus hannai (Cheng et al., 2007)와 같은 연체동물 종 일부에서 확인되고 있다. 이와 같이 HSP family 중에서도 HSP70 유전자는 환경 변화 스트레스에 민감하는 열스트레스 단백질로 알려져 있으며, 세포의 성장과 억제에 대한 빠른 분자적 반응을 나타내어 스트레스 지표유전자로서 다양한 연구가 진행 중이다.
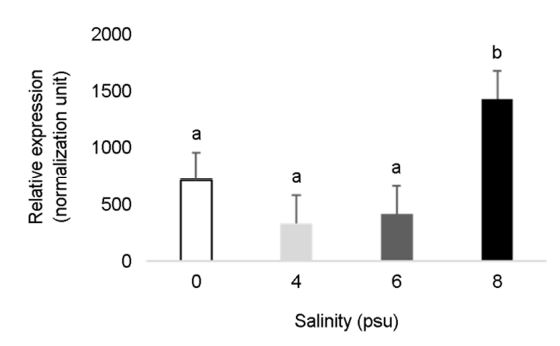
생리학적 변화를 유도하는 요인 중 하나인 염분 변화는 생물의 신진대사, 성장, 면역 등 영향을 미치는 것으로 알려져 있다(Baltzegar et al., 2013; Moshtaghi et al., 2016). 본 연구에서는 염분에 노출된 지 24시간 경과 후 10 psu 실험구를 제외한 모든 실험구에서는 폐사가 확인되지 않았다. 염분 노출로 인한 참다슬기 HSP70 발현량을 살펴보면, 대조구에 비해서 염분 8 psu 구간에 노출된 참다슬기 아가미 조직에서 유의적으로 약 2배 정도 증가된 HSP70 양을 확인할 수 있었다(Fig. 5). 해수에 노출된 꽃게 Scylla paramamosain와 해삼 Apostichopus japonicus에서도 HSP70 발현양이 증가하는 것이 보고되었다(Yang et al., 2013; Wang et al., 2014). 담수에 서식하는 깔따구 Chironomus riparius의 경우 염분 농도 5, 10 psu에 노출시켰을 때 HSP70 유전자 발현이 약 2.5배~4배 이상 증가하였다(Kim et al., 2017). 이는 환경 중의 이온이 체내로 유입됨으로써, 생리적인 불균형을 유발하고 이는 유해물질과 유사한 생태독성학적 반응을 나타내었다. 염분 유입과 같은 환경 변화는 담수에서만 서식할 수 있는 생물체 내에 해로운 영향력을 미칠 수도 있다는 사실을 반증하고 있다. 염분(8 psu)에 24시간 노출된 담수 참다슬기 개체 중에서 폐사가 확인되지 않았으나, 거머리는 동일한 염분 농도와 노출시간을 적용시켰을 때 폐사가 관찰되었다(data not shown). 이러한 결과는 염분을 이용한 친환경적인 방법을 이용하여 담수 참다슬기 양식 시 거머리를 제거할 수 있는 근거를 제공할 수 있을 것으로 예상된다. 또한 추후 더 세밀한 실험설계로 염분 변화 시 일어나는 이온 이동에 관여하는 막단백질과의 HSP70 유전자의 상관관계를 검토함으로써 수산생물의 삼투생리를 심도있게 이해할 수 있는 분자적 마커로서 활용할 수 있을 것으로 예상된다.
- References
-
1. Baltzegar DA, Reading BJ, Douros JD, Borski RJ. 2013. Role for leptin in promoting glucose mobilization during acute hyperosmotic stress in teleost fishes. J Endocrinol 220: 61-72. doi: 10.1530/JOE-13-0292.


-
2. Cellura C, Toubiana M, Parrinello N, Roch P. 2006. HSP70 gene expression in Mytilus galloprovincialis hemocytes is triggered by moderate heat shock and Vibrio anguillarum, but not by V. splendidus or Micrococcus lysodeikticus. Dev Comp Immunol 30: 984-997.
-
3. Cheng P, Liu X, Zhang G, He J. 2007. Cloning and expression analysis of a HSP70 gene from Pacific abalone (Haliotis discus hannai). Fish Shellfish Immun 22: 77-87.
-
4. Evans DH, Piermarini PM, Choe KP. 2005. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol Rev 85: 97-177.
-
5. Farcy E, Serpentini A, Fiévet B, Lebel J. 2007. Identification of cDNAs encoding HSP70 and HSP90 in the abalone Haliotis tuberculata: transcriptional induction in response to thermal stress in hemocyte primary culture. Comp Biochem Phys B 146: 540-550.
-
6. Feder ME, Hofmann GE. 1999. Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and eco- logical physiology. Annu Rev Physiol 61: 243-282.
-
7. Han G, Zhang S, Marshall DJ, Ke C, Dong Y. 2013. Metabolic energy sensors (AMPK and SIRT1), protein carbonylation, and cardiac failure as biomarkers of thermal stress in an intertidal limpet: linking energetic allocation with environmental tem- perature during aerial emersion. J Exp Biol doi: 10.1242/ jeb.084269
-
8. Hightower LE. 1991. Heat shock, stress proteins, chaperones, and proteotoxicity. Cell 66: 191-197.
-
9. Hunter S, Apweiler R, Attwood TK, Bairoch A, Bateman A, Binns D, Bork P, Das U, Daugherty L, Duquenne L. 2009. InterPro: the integrative protein signature database. Nucleic Acids Res 37: D211-D215.
-
10. Kim DH, Kim HS, Lee WO, Lee BC, Kim JH, Hong KH. 2009. In- hibitory Effect of hydrogen Peroxide against Radix auricularia in aquatic environment. In: Conference of Korean Soc Fish aquatic Science, Korea, pp 82-83 (In Korean).
-
11. Kim WS, Im BH, Hong C, Choi SW, Park KY, Kwak IS. 2017. Gene Expression of Chironomus riparius Heat Shock Protein 70 and Developmental Retardation Exposure to Salinity. Korean J Limnol 50: 305-313.
-
12. Kim YE, Hipp MS, Bracher A, Hayer-Hartl M, Ulrich Hartl F. 2013. Molecular chaperone functions in protein folding and pro- teostasis. Annu Rev Biochem 82: 323-355.
-
13. Köhler F. 2017. Against the odds of unusual mtDNA inheritance, introgressive hybridisation and phenotypic plasticity: system- atic revision of Korean freshwater gastropods (Semisulco- spiridae, Cerithioidea). Invertebr Systemat 31: 249-268.
-
14. Laursen JR, Liu H, Wu X, Yoshino TP. 1997. Heat-shock response in a molluscan cell line: characterization of the response and cloning of an inducible HSP70 cDNA. J Invertebr Pathol 70: 226-233.
-
15. Lee T, Hong HC, Kim JJ, Foighil DÓ. 2007. Phylogenetic and taxonomic incongruence involving nuclear and mitochondrial markers in Korean populations of the freshwater snail genus Semisulcospira (Cerithioidea: Pleuroceridae). Mol Phylogenet Evol 43: 386-397.
-
16. Mcattleadanmohanrao G. 1960. Salinity tolerance and oxygen consumption of the leech Hirudinaria granulosa. In Proceedings of the Indian Academy of Sciences-Section B Springer India 51: 211-218.
-
17. Morimoto RI. 1998. Regulation of the heat shock transcriptional response: cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev 12: 3788-3796.
-
18. Moshtaghi A, Rahi ML, Nguyen VT, Mather PB, Hurwood DA. 2016. A transcriptomic scan for potential candidate genes involved in osmoregulation in an obligate freshwater palae- monid prawn (Macrobrachium australiense). Peer J 4: e2520.
-
19. Piano A, Asirelli C, Caselli F, Fabbri E. 2002. Hsp70 expression in thermally stressed Ostrea edulis, a commercially important oyster in Europe. Cell Stress Chaperon 7: 250.
-
20. Piano A, Franzellitt S, Tinti F, Fabbri E. 2005. Sequencing and expression pattern of inducible heat shock gene products in the European flat oyster, Ostrea edulis. Gene 361: 119-126.
-
21. Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 3: 1101-1108.
-
22. Song JH, Kang JH. 2016. The Current Status and Tasks of Marsh snail Restocking Project based on Economic Performance Evaluation. J Kor Soc Fish Mar Edu 28: 450-455.
-
23. Song HM, Mu XD, Gu DE, Luo D, Yang YX, Xu M, Luo JR, Zhang JE, Hu YC. 2014. Molecular characteristics of the HSP70 gene and its differential expression in female and male golden apple snails (Pomacea canaliculata) under temperature stimu- lation. Cell Stress Chaperones 19: 579-589.
-
24. Srivastava P. 2002. Roles of heat-shock proteins in innate and adaptive immunity. Nat Rev Immunol 2: 185-194.
-
25. Thompson JD, Higgins DG, Gibson TJ. 1994. Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673-4680.
-
26. Trent JD. 1996. A review of acquired thermotolerance, heat-shock proteins and molecular chaperones in Archaea. FEMS Microbiol Rev 18: 249-258.
-
27. Tort L, Balasch JC, Mackenzie S. 2003. Fish immune system. A cross- roads between innate and adaptive responses. Inmunología 22: 277-286.
-
28. Wang QI, Yu SS, Qin CX, Dong SI, Dong YW. 2014. Combined effects of acute thermal and hypo-osmotic stresses on osmolality and hsp70, hsp90 and sod expression in the sea cucumber Apostichopus japonicus Selenka. Aquaculture In- ternational 22: 1149-1161.
-
29. Welch WJ, Feramisco JR. 1984. Nuclear and nucleolar localization of the 72,000-dalton heat shock protein in heat-shocked mammalian cells. J Biol Chem 259: 4501-4513.
-
30. Yang YN, Ye H, Huang H, Li S, Liu X, Zeng X, Gong J. 2013. Expression of Hsp70 in the mud crab, Scylla paramamosain in response to bacterial, osmotic, and thermal stress. Cell Stress and Chaperones 18: 475-482.
Copyright (C) 2024. The Korean Society of Marine Life Science