










JMLS 2019 June;4(1):22-28. http://jmls.or.kr Epub 2019 June 17
Copyright © 2019 by The Korean Society of Marine Life Science
Diversity of Culturable Bacteria Associated with Hard Coral from the Antarctic Ross Sea
Min Ju Kim; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea
Ha Ju Park; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea
Ui Joung Youn; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea
Joung Han Yim; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea
Se Jong Han; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea; Department of Polar Sciences, University of Science and Technology, Daejeon 34113, Korea
- Abstract
The bacterial diversity of an Antarctic hard coral, Errina fissurata, was examined by isolating bacterial colonies from crushed coral tissue and by sequencing their 16S rRNA gene. From the analyzed results, the bacteria were classified as Actinobacteria (56%), Firmicutes (35%) and Proteobacteria (9%). The thirty-four isolates were cultured in liquid media at different temperatures and their growth was assessed over time. The majority of the isolates displayed their highest growth rate at 25℃ during the first three days of cultivation, even though the coral was from a cold environment. Nevertheless, strains showing their highest growth rate at low temperatures (15℃ and 4℃) were also found. This study reports the composition of an Antarctic hard coral-associated culturable bacterial community and their growth behavior at different temperatures.
Keywords: Antarctic Ross Sea Coral Bacteria Growth rate
Correspondence to: Se Jong Han; Division of Polar Life Sciences, Korea Polar Research Institute, Incheon 21990, Korea; E-mail : hansj@kopri.re.kr
- Received
- 14 May 2019;
- Revised
- 21 May 2019;
- Accepted
- 11 June 2019.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: English,
Full Text:

Introduction
Despite being a harsh environment, Antarctica has high biodiversity (Chown et al., 2015). Over 8,300 marine species are listed on the Register of Antarctic Marine Species (De Broyer, 2019). In particular, over 4,100 species are reported as benthic species in the Southern Ocean (Clarke and Johnston, 2003). Furthermore, new species are continuously discovered in Antarctica (Janosik and Halanych, 2010). Coral reefs harbor a large part of the diversity in marine ecosystems (Forest et al., 2002). Small et al. (1998) suggested that the number of species in coral reefs is almost 3 million in pan tropic regions, suggesting that they might be the ecosystem with the highest specific diversity. However, coral reefs are facing bleaching and calcification under climate change (Cheal et al., 2017). Changes in the members of the community harbored by coral reefs might also be an indicator of global warming. The diversity of coral-associated bacterial community provides insight into the coral’s health status (Zhang et al., 2015). A coral microbial community consists of microbes that live on the surface of corals, within the tissue, and in the surrounding water (Rohwer et al., 2001). The diversity of the microbes in the Antarctic region has been investigated continuously (Antony et al., 2012; Baek et al., 2013; Ciesielski et al., 2014), including in the soft coral-associated bacteria (Webster and Bourne, 2006). However, the microbial community of the Antarctic hard coral has not been extensively studied yet. The purpose of this work was to investigate the diversity, distribution, and characteristics of the culturable bacteria present in a hard coral found in the Ross Sea.
Materials and Methods
1. Sample collection and isolation of coral-associated bacteria
The red sea coral Errina fissurata was collected from the Ross Sea during the expedition cruise of Araon (the Korean research icebreaker) (Fig. 1), which was from February 29th to March 26th, 2016; the samples were collected on March 13th at station number 6 (73°23.1187'S, 173°20.6802'E). The dredge connected to the Araon was used to lift the biological organisms, including the red coral, from the benthic zone at a depth of 330 m.

Five pieces of coral were randomly chosen, and their surfaces were washed with sterilized water. Each piece was ground using an autoclaved mortar and pestle. 4 ml of sterilized saline solution was added to the ground coral and the solution was diluted until the colonies were able to pick individually. ISP Medium 4 (BD), Marine Broth 2216 (MB) (BD), Nutrient Broth (NB) (Sigma), Potato dextrose broth (PDB) (Acumedia), R2A (MBcell), ZoBell 2216e (ZoBell, 1946), and super ZoBell (2% of glucose added to ZoBell) agar (15%) were used to culture bacteria. Plates were incubated at 15℃ for five to seven days and colonies were selected from the agar plate based on their shape and color. Colonies were named after the number of the piece of coral (1 to 5) and the order of the colony selected. Each isolate was then stored at -80℃ as a 20% (v/v) glycerol stock solution.
2. Identification of microorganisms
The identification of the isolates were performed by PCR amplification of the 16S rRNA gene using 27F (5' AGA GTT TGA TCM TGG CTC AG 3') and 1492R (5' TAC GGY TAC CTT GTT ACG ACT T 3') primers by methods described previously (Lane, 1991). For more precise results, 785F (5' GGA TTA GAT ACC CTG GTA 3') and 907R (5' CCG TCA ATT CCT TTR AGT TT 3') primers were also used for sequencing. The sequencing was done by Macrogen Korea. Using two amplicons (1,465 bp and 122 bp), sequences were aligned and matched to strains from EzBioCloud database (Yoon et al., 2017). The strain showing the highest similarity was chosen for identification.
3. Phylogenetic tree
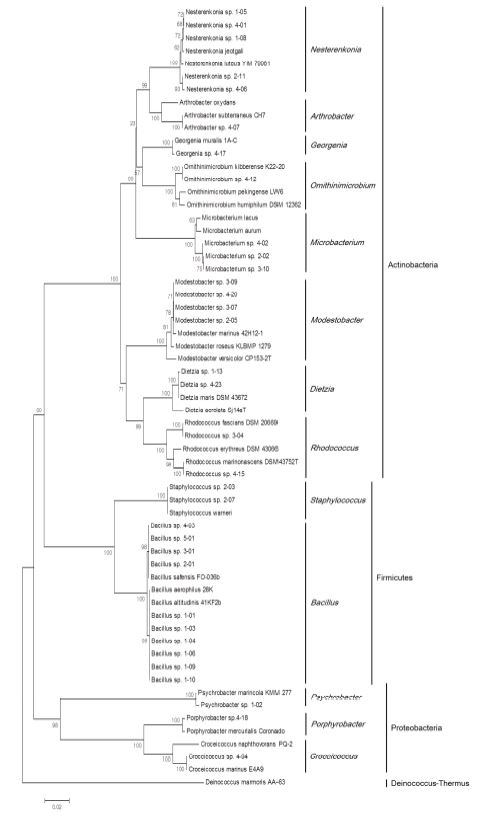
A phylogenetic tree was created based on the 16S rRNA gene sequences. Sequence information of isolates was altered to fasta file format using CLUSTAL X (Thompson et al., 1997), and sequence alignment and phylogenetic tree formation was performed by MEGA6 (Tamura et al., 2013). The alignment was performed with representative species of Actinobacteria, Firmicutes, and Proteobacteria. Deinococcus-Thermus was included as the ourgroup. The tree was constructed using the neighbor-joining method (Saitou and Nei, 1987) and calculated using Kimura's 2-parameter evolutionary model (Kimura, 1980). Bootstrap analysis to evaluate the confidence level was performed using 1,000 replications.
4. Effect of temperature on cell growth
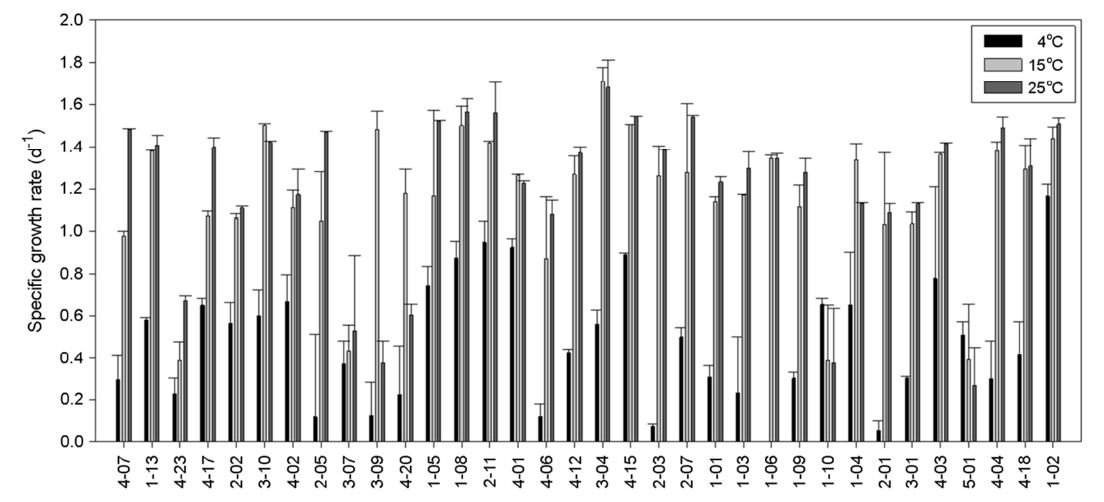
To determine the effect of temperature on cell growth, all the isolates were cultured on each media which the isolates were originally selected at three different temperatures. First, the isolates were seed cultured at 15℃, and used as the inoculum adjusting the initial cell density to OD600=0.01. Each strain was grown at 4℃, 15℃, and 25℃, and the cell density was measured on the 3rd, 7th, and 18th days after inoculation. The OD600 (optical density at 600 nm) was measured using a SCINCO S-3100 spectrophotometer (SINCO, Seoul, Republic of Korea). Specific growth rate (Berney et al., 2006) was determined at the third day of cultivation to consider the isolates' growth characteristics.
Results
1. Isolation of bacteria from an Antarctic hard coral
From the Antarctic hard coral, 34 colonies were isolated (Table 1). To obtain numerous isolates, various media was used such as ZoBell, ISP4, MB, NB, PDB, R2A, and Super ZoBell. Except Marin broth, Antarctic sea water sterilized with a membrane filter (0.22 μm pore size) was used to cultivate marine bacteria. Each media has a difference at nutrient levels such as poor to rich. Among seven media, ZoBell cultured the highest variety of isolates. The colors of isolates were yellow, beige, orange, lemon, pink, apricot, reddish, and white (Table 1). Yellow and pink pigments from Bacillus isolates are known as carotenoids and its role is to protect from light and to have more residency to UV radiation (Khaneja et al., 2010). Thirty strains (86%) had round-shaped colonies, and the others (14%) had irregular shaped colonies.

2. Identification and phylogenetic analysis
The identified isolates were classified as Actinobacteria (56%), Firmicutes (35%) and Proteobacteria (9%). Each isolate was 16 different species and 13 different genera (Fig. 2). Genus Bacillus occupied the largest portionten isolates among thirty-four. The phylum proteobacteria comprises Alphaproteobacteria (two isolates), and Gammaproteobacteria (one isolates).
3. Effect of temperature on cell growth
Sixteen isolates (47%) reached their highest density at 15℃, fourteen (41%) at 25°C, and four (12%) at 4℃ when they were seed cultured at 15℃, and then cultured at three different temperatures (Fig. 3). Even the isolates were from the harsh cold environments and capable growing at low temperatures, the isolated bacteria demonstrated better growth at higher temperatures showing psychrotolerant features (Romanovaskaia et al., 2012). However, four isolates which demonstrated highest cell growth at 4℃ were psychrophilic. Specific growth rates (μ, d-1) of each microbe were calculated. Overall, the cell growth rate of isolates at 4℃ was lower than that at the other two higher temperatures showing faster growth at high temperatures (Fig. 4). Among the 34 isolates, 25 (74%) showed highest growth rate at 25℃. This is in accord with a previous study (Ratkowsky et al., 1982). Seven isolates (21%) presented the highest growth rate at 15℃, and two isolates (1-10 and 5-01) grew faster at 4℃ than other higher temperatures. Isolate 1-10 and 5-01 seems psychrophilic about their growth.

Discussion
Bacteria are discovered in corals from diverse environments. A soft coral that inhabits the Antarctic, Alcyonium antarcticum, shows high diversity of coral-associated bacteria, including Proteobacteria, Bacteroidetes, Firmicutes, Actinomycetales, Planctomycetes, Chlorobi, and sulfate-reducing bacteria (Webster and Bourne, 2006). In contrast, the majority of Mediterranean coral-associated bacteria were disclosed to be Vibrio splendidus, which belongs to the phylum Proteobacteria (Koren and Rosenberg, 2006). Tropical corals typically cover a broad phylum of bacteria (Rohwer et al., 2001). Refer to Rohwer's study, from the tropical corals, isolate belonging to genus Bacillus was cultured which is overlapping with our results. It seems that genus Bacillus has a broad range of temperature tolerance. Here, we investigated the coral-associated cultured bacteria of Antarctic hard coral. The community comprised Actinobacteria (55%), Firmicutes (35%), and Proteobacteria (9%). Different coral species contain different microbial communities, and environmental factors affect the community (Forest et al., 2002; Zhang et al., 2015).
The temperature effect on cell growth seems relative, and the growth rate was affected by the culturing temperature. From 273.15K (0℃) to 300K (26.85℃), the cell growth rate of a Pseudomonas strain showed directly proportionate increase (Ratkowsky et al., 1982). Pseudomonas is an aerobic Gammaproteobacteria that is widespread in water environments. One of the reasons that temperature affects bacterial growth might be that temperatures below the optimum lower the enzyme affinity to substrates (Nedwell, 1999). Therefore, at low temperature, the growth of bacteria slows down. Given that most of the isolates in this study showed highest growth rate at 25℃ and a few isolates demonstrated a high growth rate at 4℃. It seems that even though the isolates were originated from the Antarctic, if a better condition such as higher temperature is given, they might expand their growth capacity.
In conclusion, we obtained 34 bacterial isolates from Antarctic hard coral. The isolates consisted of Actinobacteria, Firmicutes and Proteobacteria. Most of the isolates showed the fastest growth rate at 25℃; however, nine isolates showed a psychrophilic characteristics. To understand the diversity of hard coral-associated bacteria completely, other methods of retaining unculturable bacteria should be applied. To discover the unrevealed bacteria, genomic DNA extraction and metagenome sequencing using the coral samples should be conducted. To our knowledge, this is the first study on Antarctic hard coral-associated bacteria. Given that we used a ground sample of hard coral, we most likely isolated the bacteria located on the coral's surface and inside its tissue. Even though we only isolated culturable bacteria, we were able to reveal the diversity of the bacteria associated with Antarctic hard coral. Variations of community members might be the barometer of environmental change for the Antarctic Ross Sea.
- References
-
1. Antony R, Krishnan KP, Laluraj CM, Thamban M, Dhakephalkar PK, Engineer AS, Shivaji S. 2012. Diversity and physiology of culturable bacteria associated with a coastal Antarctic ice core. Microbiol Res 167: 372-380.


-
2. Baek K, Choi A, Kang I, Lee K, Cho JC. 2013. Kordia antarctica sp. nov., isolated from Antarctic seawater. Int J Syst Evol Microbiol 63: 3617-3622.
-
3. Berney M, Weilenmann HU, Ihssen J, Bassin C, Egli T. 2006. Specific growth rate determines the sensitivity of Escherichia coli to thermal, UVA, and solar disinfection. Appl Environ Microbiol 72: 2586-2593.
-
4. Cheal AJ, Macneil MA, Emslie MJ, Sweatman H. 2017. The threat to coral reefs from more intense cyclones under climate change. Global Change Biology 23: 1511-1524.
-
5. Chown SL, Clarke A, Fraser CI, Cary SC, Moon KL, Mcgeoch MA. 2015. The changing form of Antarctic biodiversity. Nature 522: 431-438.
-
6. Ciesielski S, Gorniak D, Mozejko J, Swiatecki A, Grzesiak J, Zdanowski M. 2014. The diversity of bacteria isolated from antarctic freshwater reservoirs possessing the ability to produce polyhydroxyalkanoates. Curr Microbiol 69: 594-603.
-
7. Clarke A, Johnston NM. 2003. Antarctic marine benthic diversity. Oceanogr Mar Biol: an Annual Review 41: 47-114.
-
8. De Broyer C, Clarke A, Koubbi P, Pakhomov E, Scott F, Vanden Berghe E, Danis B. (Eds.) (2019). Register of Antarctic Marine Species. Accessed at http://www.marinespecies.org/rams on 2019-01-21.
-
9. Forest R, Victor S, Farooq A, Nancy K. 2002. Diversity and distribution of coral-associated bacteria. Mar Ecol Prog Ser 243: 1-10.
-
10. Janosik AM, Halanych KM. 2010. Unrecognized Antarctic biodiversity: a case study of the genus Odontaster (Odontasteridae; Asteroidea). Integr Comp Biol 50: 981-992.
-
11. Khaneja R, Perez-Fons L, Fakhry S, Baccigalupi L, Steiger S, To E, Sandmann G, Dong TC, Ricca E, Fraser PD, Cutting SM. 2010. Carotenoids found in Bacillus. J Appl Microbiol 108: 1889 -1902.
-
12. Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol 16: 111-120.
-
13. Koren O, Rosenberg E. 2006. Bacteria associated with mucus and tissues of the coral Oculina patagonica in summer and winter. Appl Environ Microbiol 72: 5254-5259.
-
14. Lane DJ. 1991. 16S/23S rRNA sequencing. In E. Stackebrandt and M. Goodfellow (ed.), Nucleic acid techniques in bacterial systematics. John Wiley & Sons, New York, pp 115-175.
-
15. Nedwell DB. 1999. Effect of low temperature on microbial growth: lowered affinity for substrates limits growth at low temperature. FEMS Microbiol Ecol 30: 101-111.
-
16. Ratkowsky DA, Olley J, Mc Meekin TA, Ball A. 1982. Relationship between temperature and growth rate of bacterial cultures. J Bacteriol 149: 1-5.
-
17. Rohwer F, Breitbart M, Jara J, Azam F, Knowlton N. 2001. Diversity of bacteria associated with the Caribbean coral Montastraea franksi. Coral reefs 20: 85-91.
-
18. Romanovaskaia VA, Tashirev AB, Gladka GB, Tashireva AA. 2012. Temperature range for growth of the Antarctic microorganisms. Mikrobiol Z 74: 13-9.
-
19. Saitou N, Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425.
-
20. Small AM, Adey WH, Spoon D. 1998. Are current estimates of coral reef biodiversity too low? The view through the window of a microcosm. Atoll Research Bulletin 458: 1-20.
-
21. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30: 2725-2729.
-
22. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgin DG. 1997. The CLUSTAL X window interface: Flexible strategies for multiple sequence alignment aided by quality analysis tool. Nucleic Acids Res 24: 4876-4882.
-
23. Webster NS, Bourne D. 2006. Bacterial community structure associated with the Antarctic soft coral, Alcyonium antarcticum. FEMS Microbiol Ecol 59: 81-94.
-
24. Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, Chun J. 2017. Introducing EzBioCloud: A taxonomically united database of 16S rRNA and whole genome assemblies. Int J Syst Evol Microbiol 67: 1613-1617.
-
25. Zhang YY, Ling J, Yang QS, Wang YS, Sun CC, Sun HY, Feng JB, Jiang YF, Zhang YZ, Wu ML, Dong JD. 2015. The diversity of coral associated bacteria and the environmental factors affect their community variation. Ecotoxicology 24: 1467-1477.
-
26. Zobell CE. 1946. Marine Microbiology. A Monogram on Hydro-bacteriology. Chronica Botanica Co., Waltham, 240 p.
Copyright (C) 2024. The Korean Society of Marine Life Science