










JMLS 2019 June;4(1):14-21. http://jmls.or.kr Epub 2019 June 17
Copyright © 2019 by The Korean Society of Marine Life Science
Review of Migration and Distribution of the Common Squid (Todarodes pacificus) in the East Sea and the Yellow Sea
Yoon Ha Kim; Gangneung-Wonju National University, Gangneung 25457, Korea
Hae Kun Jung; Gangneung-Wonju National University, Gangneung 25457, Korea
Sung-Yong Oh; Korea Institute of Ocean Science & Technology, Busan 49111, Korea
Hyun Woo Kim; Pukyung National University, Busan 48513, Korea
Chung Il Lee; Gangneung-Wonju National University, Gangneung 25457, Korea
- Abstract
본 리뷰는 한국의 중요한 수산자원인 살오징어(Todarodes pacificus)의 동해와 서해 어획량 변동원인을 살오징어 난/자치어 수송부터 회유경로 및 어장 분포에 관점으로 논의하였다. 우리나라 살오징어 어획량은 1980년 이후 기후체제전환에 따라 변화가 있어 왔으며 이는 동해와 서해의 어획량 변동 경향이 달랐다. PDO (Pacific Decadal Oscillation)는 동해로 유입되는 난류 수송량과 음의 상관관계가 있는데 PDO가 양의 위상이었던 1980s에는 서해에서는 어획량이 많았고 동해에서는 어획량이 적었다. 반면, PDO가 음의 위상이었던 1990s년대에는 동해 어획량이 많고 서해에서는 적었다. 이는 살오징어가 난류를 따라 북상하거나 난류를 거슬러 남하회유를 하는 생활사에 기인된다. 동해의 경우, 난류가 강(약)할 때, 난류경로가 한국의 동해 연안 쪽(동해 중부 해역 및 일본 연안 쪽)으로 치우치게 되는데 이는 PDO가 양의 위상이었던 1980년대에 어장이 울릉도 동편에 위치하였던 반면 PDO가 음의 위상이었던 1990년대에 어장이 동해 연안에 위치한 것과 관련있다. 서해 살오징어 어획량이 증가한 1980년대에는 동해로 유입되는 난류수 수송량이 감소한 반면 서해로 유입되는 난류수는 증가하였으며 이는 서해로 수송되는 유생의 양을 증가시키는 주요 원인이 된다.
This review paper discussed the decadal fluctuations in the catch of the common squid, Todarodes pacificus (T. pacificus) by focusing on migration and distribution patterns. Since 1980s, changes in T. pacificus catches were due to climate regime shift in Korean waters. Fluctuation patterns of catches were different between the East Sea and the Yellow Sea. Generally PDO (Pacific Decadal Oscillation) phase shows a negative correlation with strength of warm current to the East Sea. In 1980s when PDO was positive phase (+), T. pacificus catch was higher in the Yellow, but it was lower in the East Sea. In 1990s when PDO was negative phase (-), T. pacificus catch showed opposite trend compared with 1980s. Such spatial and decadal fluctuations of T. pacificus catch were due to its northward migration along with the warm current or southward movement against the current. In the East Sea, strong (weak) warm current period, the current path has been shifted toward the East Sea coast of Korea (central East Sea or the coast of Japan). It has a correlation with PDO. In the positive PDO phase (1980s), the fishing ground was located on the eastern side of Ulleungdo, whereas during negative PDO phase (1990s), they were situated near the southeastern coast of the Korean peninsula. In the 1980s, volume transport passing into the Yellow Sea increased, whereas volume transport in the East Sea decreased. This is one of major reason increasing T. pacificus larvae in the Yellow Sea.
Keywords: Todarodes pacificus Migration(회유) Distribution(분포) East Sea(동해) Yellow Sea(서해) Pacific Decadal Oscillation(태평양순년변동)
Correspondence to: Chung Il Lee; Gangneung-Wonju National University, Gangneung 25457, Korea; E-mail : leeci@gwnu.ac.kr
- Received
- 17 May 2019;
- Revised
- 27 May 2019;
- Accepted
- 4 June 2019.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
살오징어, Todarodes pacificus는 연체동물문(Phylum Mollusca)-두족강(Class Cephalopoda)-개안목(Order Oegopsida)-살오징어과(Family Ommastrephidae)에 속한다(Roper et al., 1984). 산란장과 색이 · 성육장을 생애에 걸쳐 회유하며, 대만 근해로부터 동중국해 및 일본 태평양측 연안과 오호츠크 해까지 널리 분포한다(Okutani, 1983; Roper et al., 1984; Murata, 1990; Shevtsov et al., 2005). 산란시기에 따라 하계군, 추계군, 동계군의 산란계군으로 구분되며, 각 계군마다 차이는 있으나 주산란장은 동중국해-큐슈 서안을 중심으로 형성된다(Okutani, 1983; Murata, 1990; Sakurai et al., 2000; Sakurai et al., 2002). 산란장에서 난, 유생 및 어린 살오징어는 난류를 따라 북상하여 동해 북부 및 오호츠크 해로 수송되어 성장하며, 이후 산란을 위해 남하회유를 한다(Okutani, 1983; Roper et al., 1984). 살오징어 어장의 시/공간적인 분포 변화에 영향을 미치는 가장 중요한 인자는 수온이다. 살오징어는 광온성으로 알려져 있나 어획적수온은 17℃를 중심으로 한 15~19℃의 범위이다(Lee et al., 1985). 어장은 주로 한류와 난류가 만나는 전선의 시/공간적인 분포 변화에 따라 형성되며, 수온의 분포가 어황에 영향을 미친다(Choi, 2005; Choi et al., 1997, 2003; Mokrin et al., 2002; Cho et al., 2004; Kim et al., 2010, 2017, 2018). 북서태평양에서 주요 살오징어 어장은 우리나라 동해와 일본의 동쪽(태평양 측)이 중심이 되어 형성되며, 주 어획국가는 한국과 일본이다. 식량농업기구(Food and Agriculture Organization: FAO)에 따르면 살오징어 어획량은 수년에서 십수년 주기로 변동이 있었다(Fig. 1).
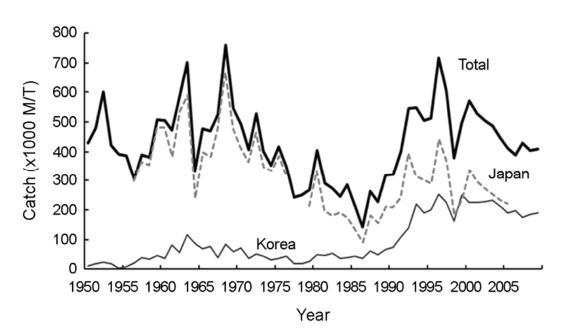
이러한 살오징어 어획량의 변화는 어획노력과 같은 인간의 인위적인 요인과 자연환경 변화에 의한 요인을 동시에 받는다(Kim et al., 2010, 2017, 2018). 근대적인 항법체계, 냉동독항선 그리고 자동조상기 등의 조업기술발달 및 경제성에 따라 어획노력이 증가하여 이에 따라 어업강도가 변화하였으며(Murata, 1990; Choi, 2005), 이러한 어업강도의 변화는 친어 자원량의 변화로 작용하여 산란 자원량 및 가입량에 영향을 주어 결국 어획량에 영향을 미칠 수 있다. 후자는 수온 변화와 그에 따른 생물학적 반응과 같은 해양환경 변화에 따른 산란시기, 어장 분포 및 회유경로의 변화이다.
Sakurai et al. (2000)은 기후체제 전환기와 살오징어 어획량의 급변한 시기가 연관이 있다고 보았다. 기후체제전환은 장기간의 평균적인 대기와 해양의 상태 변화를 의미하는데 지난 세기 후반에 기후체제전환은 3차례가 있었으며(Minobe, 1997; Francis et al., 1998; Mantua and Hare, 2002; Hare and Mantua, 2000; Sakurai et al., 2000, 2002; Overland et al., 2008), 온난기였던 1970s, 1990s에는 우리나라 동해를 포함한 북서태평양의 살오징어 어획량이 증가하였으며, 반대로 한랭기였던 1980s에는 감소하였다(Sakurai et al., 2000; Kim, 2015).
기후체제전환에 따른 해양환경 변화는 산란장 환경 변화를 야기하여 살오징어 산란량, 유생의 생존량 등에 영향을 미칠 수 있으며, 이것이 가입량에 변화를 주게 되어 어획량 변화에까지 그 영향이 전달된다(Sakurai, 2006; Sakurai et al., 2000, 2002; Kim et al., 2017, 2018). 이는 1980년대 후반부터 북서태평양 살오징어 어획량 증가의 주요 원인이라 할 수 있다. 그러나 우리나라 살오징어 어획량의 경우, 급증한 주요 원인을 자원량 변화만으로는 해역별 어획량 변동을 설명하기 어렵다.
같은 동해일지라도 한국 동안(동해 서편)의 살오징어 어획량은 1980년대 후반 이후 급증한 반면 일본 서안(동해 동편)의 어획량 변동이 현저히 작았다(Kidokoro, 2009). 서해에서는 북서태평양의 어획량이 적었던 1980년대에 오히려 더 많은 경향을 보였다(Kim, 2015). 이는 어획에는 자원량 뿐만 아니라 어장환경 또한 반드시 고려되어야 함을 의미한다.
살오징어 어장환경 변화의 주요 인자는 수온으로 어획적수온은 15~19℃이며(Lee et al., 1985), 이는 산란을 위해 남하회유하는 살오징어 난소가 성숙하는 수온 15~18℃ (Kidokoro and Sakurai, 2008)와도 거의 일치한다. 따라서 수온전선의 시공간적인 분포 변화는 산란을 위해 남하하는 살오징어 회유경로 변화에 영향을 미치는 주요 인자로 작용하며, 이는 어장의 위치 및 어군의 밀도 변화에까지 그 영향이 전달된다(Cho et al., 2004; Choi, 2005; Choi et al., 1997, 2003, 2008; Kim et al., 2010, 2017; Mokrin et al., 2002).
대한해협을 통해 동해로 유입되는 쓰시마 난류는 대기-해양순환 변화에 의해 그 강약이 달라지고 그에 따라 살오징어의 회유경로가 변화한다(Hong and Cho, 1983; Lee, 2003). 쓰시마 난류 흐름의 강한 시기에는 해류경로 쓰시마 난류의 중심축이 한국 동해안으로 인접하나 약한 시기에는 동해 중앙부 또는 동해 일본 연안에 치우친다. 이러한 쓰시마 난류수 중심축의 변화는 살오징어 회유 및 분포의 변화를 일으켜 어장 위치를 변화시키기도 하는데, 한랭기와 온난기에 쓰시마 난류 분포역이 달라지며, 이에 따라 동해 북부에서 남하하는 살오징어 회유경로가 한랭기의 울릉도 주변 해역을 비롯한 동해 중부 해역 및 일본 연안에서 온난기의 동해 연안으로 변화한다(Kidokoro et al., 2010; Kim et al., 2017).
난류의 변화는 전지구규모의 대기-해양순환 변화에 영향을 받는다. 특히 태평양 20°N의 표층수온을 지수화한 Pacific Decadal Oscillation(이하 PDO)는 동해로의 난류 유입량과 음의 상관성이 있을 뿐만 아니라 서해 살오징어 어획량 변화와 유의한 상관성을 보인다(Miller et al., 2004; Jung, 2013; Kim, 2015). 이는 기후-해양환경 변화가 살오징어 어장형성에 영향을 미치는 회유 및 분포역의 변화와도 밀접한 연관이 있다는 것을 의미한다.
따라서 본 리뷰는 살오징어 어장형성원인과 변화를 분석한 기존 연구들을 토대로 하여 기후-해양환경 변화가 살오징어의 어장형성 변화에 미치는 영향을 통해 회유 및 분포의 변화특성을 논의하였다.
회유 및 분포
살오징어의 난은 다른 두족류에 비해 작고(0.8×0.7 mm) 많은 수(약 200,000립)의 알을 낳으며 짧은 부화기(4~6일)를 거쳐 작은 크기의 유생(0.95 mm)으로 발생한다(Bower and Sakurai, 1996). 산란장에서 성육 · 색이장으로의 해류에 의한 수송을 통하여 유생을 효율적으로 분산시키는 효율적인 생식전략으로 볼 수 있다(Watanabe et al., 1996). 특히나 쿠로시오와 쓰시마 난류가 지나는 경로에 산란장이 위치해 있기 때문에 난, 유생 및 미성어는 해류에 의해 수송이 일어난다.
한국 동해 남부 해역으로 수송된 유생은 난류역을 중심으로 분포하고 있다(Kim et al., 2014; Kim and Lee, 2016; Kim and Shin, 2019). 큐슈 부근의 쿠로시오역에서 채집된 유생이 쿠로시오와 큐슈 연안수가 만나는 전선역에서 밀도가 가장 높았다는 사실(Bower et al., 1999)로 미루어 보아 난류에 의해 수송되는 유생은 서로 다른 수괴가 만나는 해역에서 주로 출현하는 것으로 추정된다.
동해 남부 오키 섬 근처에서 채집된 0.7~7.3 mm 외투장분포범위의 유생수직분포경향은 75 m보다 얕은 수심부터 나타나 표층에서 25 m 범위에 현존량이 집중되었으며, 큰 개체일수록 더 깊은 수심에 분포한다(Yamamoto et al., 2007). 그리고 유생기를 지나 5 cm 정도가 되면 성체와 형태학적으로 유사하고 약한 유영능력을 지니며(Murata, 1990), 성장할수록 유영능력이 향상된다. 성어의 경우, 동해에서 채낚기 조사 시 어획된 개체의 체장과 채집된 수심이 양의 상관관계가 나타났다(Lee et al., 1985).
이처럼 유생 혹은 10 cm보다 작은 어린 개체는 주로 표층수온 15℃ 이상 수온대에 주로 분포하는 반면, 15 cm보다 큰 개체는 차고 깊은 수층에 분포하며, 전선역의 냉수역 부근에서 높은 밀도의 어군이 형성된다(Kasahara, 1978; Sakurai et al., 2000; Kawabata et al., 2006). 이와 같이 살오징어는 체장이 커짐에 따라 낮은 수온대의 깊은 수심에서 서식하는데 그 이유에 대해서는 명확히 알려져 있지 않으며, 앞으로 이에 대한 연구가 추가로 진행되어야 할 필요가 있다.
살오징어의 서식수온과는 별개로 어획적수온은 17℃를 중심으로 한 15~19℃이다(Lee et al., 1985). 어획은 주로 동해에서 동한 난류와 북한 한류, 태평양에서는 쿠로시오와 오야시오가 만나는 전선역에서 일어나기 때문에 해류의 시공간적인 분포가 중요하다. 해류 분포 양상에 따라 수평적으로는 전선의 형성방향에 의해 어군밀집 정도가 달라졌으며, 수직적으로는 수온약층의 깊이 차이로 인해 고위도로 갈수록 어획적수심은 얕아졌다(Cho et al., 2004; Choi, 2005; Choi et al., 1997, 2003, 2008; Kim et al., 2010; Mokrin et al., 2002).
북서태평양의 살오징어 어획량은 수년 주기와 십수년 주기 변동이 있었고 그 주기의 장기적인 양상은 기후체제 전환기와 일치했다(Sakurai et al., 2000). 북서태평양에서 살오징어 어획량은 저수온기였던 70년대 후반에서 80년대 중후반에 낮았던 반면 고수온기로 전환이 일어난 90년대에 접어들어 급증하였다. 어획량 변화 양상은 차이가 있으나 한국에서는 어획량이 적었던 80년대와 어획량이 많았던 1990-2000년대로 증감 양상이 유사한 반면, 일본 동해 측에서는 급격한 증감은 없었고 오히려 90년대에 비해 80년대에 높았다(Fig. 2). 이는 살오징어 자원량 증감이 절대적으로 살오징어 어획량 증감에 영향을 미치는 것이 아니라는 것을 보여준다. 따라서 어획량에 대한 해석은 자원량뿐만 아니라 해역적인 특성도 함께 고려되어야 한다.
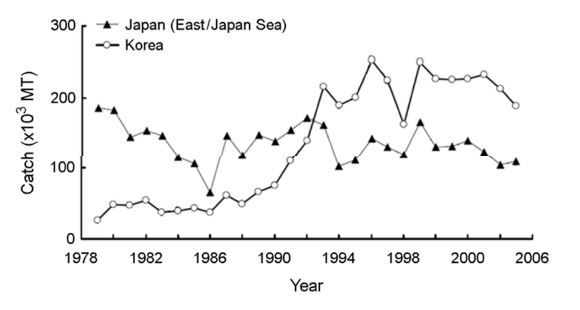
한국의 경우는 저수온기였던 80년대에는 서해 어획량이 동해보다 높았으며, 동해에서는 남동부 연안 CPUE가 높았고, 울릉도 및 외해에 총어획량이 높았다. 반면 고수온기였던 90년대에는 서해에서 살오징어가 거의 잡히지 않았고, 동해에서는 남동부 연안에서 총어획량이 높게 나타났고 울릉도 및 외해에 CPUE가 높게 나타났다(Kim et al., 2010).
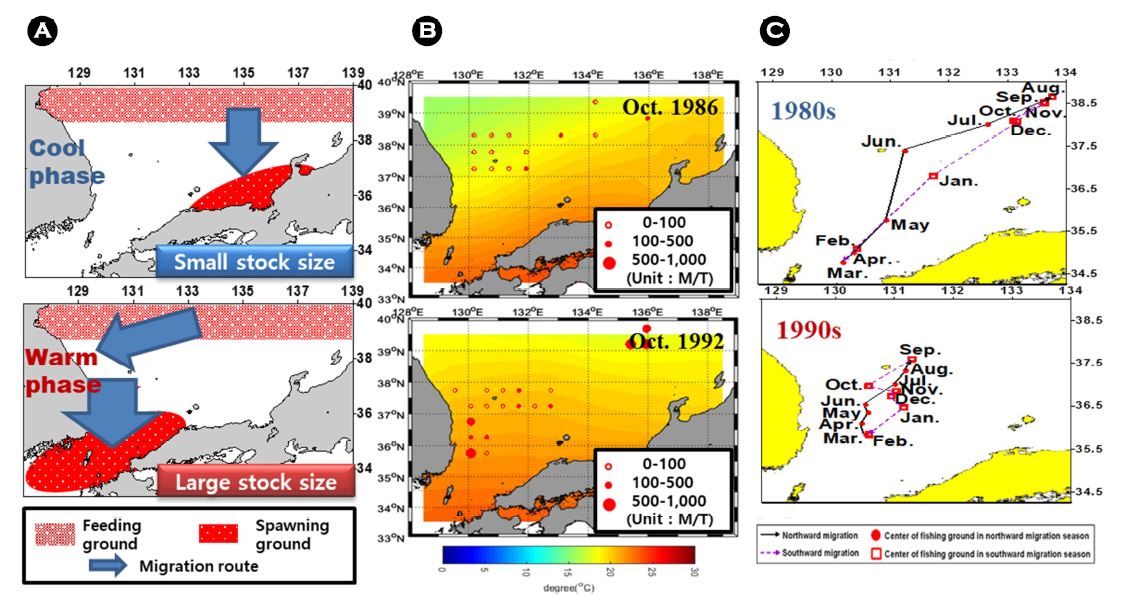
동해의 살오징어 어획량이 낮았던 해(예를 들어 1986년)는 동해의 외해(동해 중앙부)에서 살오징어 어획이 주로 이루어졌던 반면 어획량이 높았던 해(예를 들어 1992년)는 포항과 울릉도 사이의 해구에서 평년에 비해 그 어획량이 높았다(Kim et al., 2010). 어획량이 차이가 나는 두 시기(1986년, 1992년)의 10월 수온수평분포를 살펴보면 시기별로 어획량이 높았던 해역과 어획적수온대가 형성된 곳이 거의 일치함을 보였는데(Fig. 3(B)), 이는 쓰시마 난류의 세력 변화와 관계 깊다. 동해 상층부의 수온은 쓰시마 난류의 영향을 크게 받는데, 이것은 쓰시마 난류의 강약에 따라 쓰시마 난류 이동경로가 변화하며, 쓰시마 난류가 강할 때는 대한해협 통과 후 우리나라 동해안을 따라 북상하려는 경향이 강하지만, 그 세력이 약한 경우는 일본 연안을 따라 흘러가는 경향을 보인다(Hong and Cho, 1983; Lee, 2003).
따라서 이러한 살오징어의 분포 및 회유경로 변화에는 쓰시마 난류 유로 변화가 가장 큰 요인이라 생각된다. 이는 고수온기에는 계군이 한국 동해안 쪽으로 이동하여 산란하기 때문에 한반도와 대한해협 쪽으로 산란장이 이동하는 경향이 있고, 저수온기에는 동해 중앙으로 이동하는 경향으로 인해 산란장이 혼슈의 중앙 쪽으로 이동하는 경향이 있다는 Kidokoro et al. (2010)의 결론과도 일치한다(Fig. 3 (A)). 또한 이러한 분포 경향을 보이는 원인으로는 살오징어의 수온에 대한 생리적 특성에 의한 것으로 추정된다. Kidokoro and Sakurai (2008)는 1999년 동해 남부와 동중국해에서 조사된 살오징어의 생식소 발달과 수온과의 관계를 조사하였는데, SST가 15℃부터 암컷의 생식소가 발달되고 특히 SST가 15~18℃일 때 성성숙도가 최대였던 반면 외투장근육량이 낮아지는 쇠약현상이 관찰되었다. 어획 가능한 살오징어는 성숙상태 혹은 비산란기의 성체를 나타내며, 14~19℃의 어획적수온(Lee et al., 1985)의 의미는 어획 가능한 개체가 가장 빈번히 잡히는 수온이라는 것을 나타낸다. 결론적으로 동해에서 살오징어는 14~19℃ 수온대의 쓰시마 난류를 거스르며 산란장으로 남하회유한다고 추정할 수 있다.
이러한 생물학적 특성에 따라 난류의 유로 변화는 살오징어 회유경로를 변화시켜 어군이 밀집하는 장소를 변화시킨다는 것을 의미한다. 따라서 이러한 기작에 의해 어군밀집장소의 변화가 어획량에 영향을 미쳤다고 할 수 있다. 동해의 월별 어장 분포로부터 어장중심을 계산하여 기후체제별로 나타낸 결과(Fig. 3 (C)), 1980년대에는 어장중심사이의 거리가 멀었으며, 울릉도 외측에 주로 어장중심이 위치해 있었던 반면 1990년대에는 어장중심간 거리가 짧고 동해 연안 쪽에 어장중심이 위치했다(Kim et al., 2017). 이는 난류 변화가 수온 변화에 영향을 주며 이에 따라 수온에 민감하게 반응하는 살오징어의 회유경로 변화에 영향을 미쳤기 때문으로 판단된다.
서해에서는 1981년부터 형성된 어장이 1989년까지 지속되었으며, 한국 근해에서 살오징어 어획량은 1990년대에 비해 전체적으로 적었으나 서해에서 어획비율은 1981년부터 40% 전후 어획비율을 보였으며 최대어획기인 1987년에는 약 80%에 달하였다(Fig. 4). 이를 Choi et al. (2003)은 전선의 분포방향으로 인한 어군밀집조건형성 유무가 주요한 원인이라 보았다. 현재까지 1980-1990년대의 동해와 서해의 상반된 어획량변동원인에 대한 연구는 전무한 상태이다. 최근의 연구결과들로서 두 해역에서 어획량 변동 경향이 반대로 나타나는 원인은 두 가지로 추측되고 있다(Kim et al., 2017, 2018).
첫 번째 원인으로는 어업경제성 측면에서 볼 수 있다. 1980년대에 연안에서 어획량이 적었던 반면 울릉도를 중심으로 한 동해 중앙부에서 어장이 형성되었다. 따라서 먼바다로 나가 조업을 하는 것은 수익성이 낮기 때문에 비교적 어업수익성이 높았던 서해에서 주로 조업이 이루어졌다고 추정된다. 특히, 1980년대 후반 서해의 전체 위판 어획량보다 채낚기 어업 어획량이 더 많았는데(Kim, 2015), 위판자료는 채낚기 어업을 비롯한 모든 어업의 어획량을 위판되는 지역을 기반으로 나타낸 자료인 반면 해구별채낚기 어획량 자료는 어장기반자료로서, 어획되는 위치와 판매되는 위치가 다른 것은 다른 해역에서 온 어선세력이 서해에서 조업을 했다는 것의 근거로 볼 수 있다. 그러나 1990년에 접어들면서 해류의 경로가 변화하면서 동해의 어장이 연안 가까이 형성되면서 서해에서 어획하는 것보다 동해에 회유하는 자원량이 많아 어획하는데 있어 더 수익성이 높았기 때문에 동서해의 어획량 차이가 생긴 것으로 추정된다(Kim, 2015; Kim et al., 2010). 이는 동해와 서해에서 조업한 20년 동안(1980-1999년) 채낚기어선 당 어획량(CPU: catch per a vessel a day)으로 뒷받침된다. 1980년대에는 동해보다 서해가 비교적 높은 조업어선 당 어획량을 보였으나 서해에는 1987년을 기점으로 감소하는 경향을 보인 반면, 동해에서는 1980년대 후반에 접어들어 증가추세를 보이다가 1990년대 중후반에 낮아지는 경향을 보였다(Fig. 4).
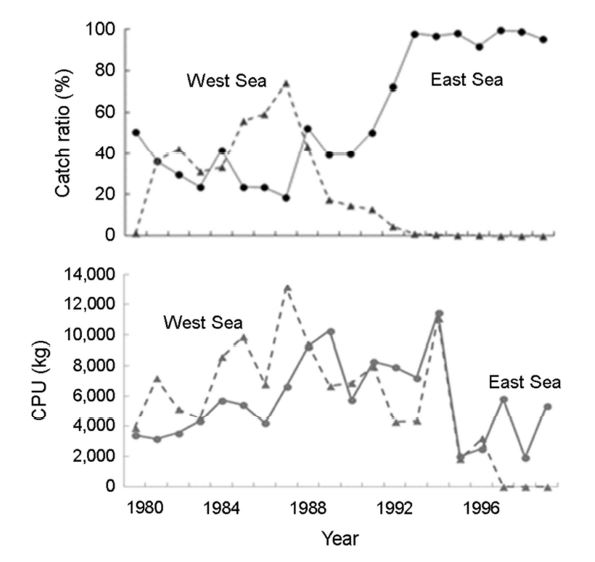
결론적으로 어선 한 척당 효율이 1980년대 후반까지는 서해에서, 그리고 1980년대 후반부터는 동해에서 더 좋았다는 것을 시사하며, 이는 1980년대 후반까지는 서해가 더 어획하기 좋은 해역이었으며, 1980년대 후반 이후로는 동해에서 어획하기에 더 좋은 해역이었음을 의미한다.
두 번째 원인으로 산란장의 규모 및 위치 및 해류 변화에 따라 달라지는 서해로 유입되는 유생 자원량의 변화를 들 수 있다. 산란장 면적과 어획량 변동의 장기적인 증감 경향이 유사하며, 산란장 크기와 위치는 기후체제전환에 의해 민감하게 변화하며 자원량에 영향을 미치고 이에 따라 어획량에 영향을 받는다(Sakurai et al., 2000; Sakurai et al., 2002; Sakurai, 2006; Rosa et al., 2011). 서해의 경우, 그 어획량은 1980년대 중후반처럼 동해를 추월한 경우도 있었고, 2000년대 중반처럼 평년에 비해 어획비율이 높아진 때도 있었다. 이는 유생의 유입량 변화에 의해 어획량이 변화했을 것으로 추정되고 있다.
Kim (2015)은 이에 대해 서해 살오징어 어획량이 PDO (Pacific Decadal Oscillation)와 상관성이 있다는 것에 주목하였다. PDO는 태평양 20°N 북측의 월별표면수온편차(SST anomaly)의 경험적 직교함수에 대한 시간계수이며(Mantua et al., 1997), 한랭기였던 1980s에는 북서태평양에서 PDO는 양의 위상으로 수온이 낮았던 반면 온난기인 1990s, 2000s에는 PDO가 음의 위상으로 이전 기후체제에 비해 수온이 높았다.
PDO가 양의 위상이었던 1980s에는 서해 어획량이 많았던 반면, PDO가 음의 위상이었던 1990s년대에는 서해 어획량이 적었다. PDO는 동해로 유입되는 난류 수송량과 음의 상관관계를 나타낸다(Jung, 2013). 즉, PDO가 양의 위상일 때, 동해로의 난류 유입량이 줄어 서해로의 유입이 많아지게 되며, 이에 따라 서해로 수송되는 유생이 증가할 가능성이 높다. 또한 계절적으로는 동계의 쓰시마 난류가 추계보다 약하기 때문에(Pang and Oh, 2000), 동계 쓰시마 난류 세력권이 제주도 서편까지 확대되어 서해로의 유생 유입 확률이 늘어나게 된다. Choi (2005)에 따르면 서해에 추계군보다 동계군의 비율이 높은 것은 이 때문으로 판단된다. 특히 PDO는 이듬해 서해 어획량과 더 높은 상관관계를 보이기 때문에 서해로 수송되는 유생의 연간 유입량의 변화는 PDO와 높은 상관성이 있다고 생각된다(Kim, 2015).
지금까지 기술한 바와 같이 살오징어는 자원량 및 어획량이 자연환경적인 요인에 의해 지속적으로 변동해왔다. 또한 앞으로도 기후는 지속적으로 변화할 것이며, 해양 또한 기후시스템의 일부로서 대기와의 상호관계를 이루며 변화가 일어날 것이다. IPCC (Intergovernmental Panel on Climate Change)에서는 기후 변화 및 그 예측에 관한 보고서를 내놓고 있는데, 2007년 Assessment Report 4 (AR4)에서 IPCC SRES (Special Report on Emission Scenarios) 시나리오의 A1B(빠른 경제성장, 세기중반에 최고인구, 빠른 새롭고 유용한 기술발전의 가정하에 화석연료를 적절히 사용할 시)에 따르면 지구의 표면온도가 지난 세기에 비해 21세기 후반에 1.7~4.4℃의 상승이 예상된다.
시나리오 중 A1B를 모델 MICRO_hire와 MPI_echam5를 통해 1990년대와 2030년대의 살오징어 산란장을 비교 분석하였을 때, 추계군과 동계군 모두 수온상승으로 인해 산란장의 북상이 예상되며(Kim, 2012), 이에 따라 산란장의 면적 변화 뿐만 아니라 회유경로, 분포까지 변화할 것으로 추측된다. 기후 변화에 따른 살오징어 어장 변화에 대한 예측이 필요하다. 이를 위해서는 정확한 예측을 위해 더욱 정교한 기후 모델과 생태계 모델이 요구되며, 이를 통해 정밀한 자원평가 및 중장기적인 어획량 예측에 도움을 줄 것으로 기대된다.
결 론
지금까지 살오징어 어획량이 변화한 원인을 기후 변화 및 해양환경 변화가 회유 및 분포에 미친 영향의 관점으로 서술하였다. 일차적으로 자원량은 기후 변화에 따른 초기생활사에 영향을 받는다. 이후에는 어장 및 어획량은 해류에 지배적인 영향아래 놓이게 되는데 살오징어 어획량에 영향을 주는 적도 발원의 쿠로시오와 그 지류인 쓰시마 난류는 PDO와 연관이 있다. PDO는 수온뿐만 아니라 해류 수송량과도 상관이 있다(Gordon and Giulivi, 2004; Jung, 2013).
살오징어 어장은 난류(warm water)의 분포 및 이동경로와 밀접한 관련이 있는데, 이러한 환경 요인은 각 국의 어획량 변화에도 영향을 끼치게 된다. 예를 들어, 한랭기(cold regime)였던 1980년대에는 쓰시마 난류가 일본 쪽으로 치우치는 경향이 있었는데 이 시기에는 연안 쪽보다 외양 쪽에 어획량이 더 높았으며, 온난기(warm regime)였던 1990년대에는 쓰시마 난류가 한국의 동해안 쪽으로 치우쳐 흘렀는데 이 시기에는 동해 남동부 연안에서 어획량이 높게 나타났다.
서해에서는 PDO와 살오징어 어획 변동량에는 양의 상관관계가 있으며, 이는 동해의 난류 수송량 변화에 따라 서해로 난류 유입량이 변화하는데 이에 따라 난류에 의해 서해로 수송되는 살오징어 유생량이 변화되기 때문이다(Kim, 2015).
그러나 PDO가 살오징어 어획량에 미치는 영향에 대한 정량적인 분석단계에는 아직 도달하지 못하고 있다. 따라서 기후, 해양생태계 그리고 수산자원의 관련성에 대한 연구를 위해 대기 및 해양의 기상자료에 근거한 기후지수 및 기후지수들 간의 정밀한 분석이 요구된다
또한 이와 같이 살오징어 어황은 단기적인 환경 변화뿐만 아니라 기후 변화와 같은 장기적인 환경 변화에 의해서도 분포와 자원량의 변화가 크게 나타난다. 따라서 지속 가능한 이용을 위해서는 기후 변화에 따른 해양환경과 살오징어 어장 변화 경향을 전망하고 시나리오별 대응방안 수립이 필요하다.
- References
-
1. Bower JR, Sakurai Y. 1996. Laboratory observations on Todarodes pacificus (Cephalopoda: Ommastrephidae) egg masses. Ame Malacol Bull 13: 65-71.


-
2. Bower JR, Nakamura Y, Mori K, Yamamoto J, Isoda Y, Sakurai Y. 1999. Distribution of Todarodes pacificus (Cephalopoda: Ommastrephidae) paralarvae near the Kuroshio off southern Kyushu. Jap Mar Biol 135: 99-106.
-
3. Cho KD, Kim SW, Kang KH, Lee CI, Kim DS, Choi YS, Choi KH. 2004. Relationship between fishing condition of common squid and oceanic condition in the East Sea. J Kor Soc Mar Env Safety 10: 61-67.
-
4. Choi KH. 2005. Fishing conditions of common squid, Todarodes pacificus (STEENSTRUP) in relation to oceanic conditions in Korean waters. Ph. D. Dissertation, Pukyung National University, Busan, Korea.
-
5. Choi KH, Cho KD, Kim DS, Kim JI, Kim SW. 2003. Fishing conditions of common squid (Todarodes pacificus Steenstrup) in the Yellow Sea. J Kor Fish Soc 6: 21-31.
-
6. Choi KH, Hwang SD, Kim JI. 1997. Fishing conditions of common squid (Todarodes pacificus STEENSTRUP) in Korean Waters. J Kor Fish Soc 30: 513-522.
-
7. Choi KH, Lee CI, Hwang KS, Kim SW, Park JH, Gong Y. 2008. Distribution and migration of Japanese common squid, Todarodes pacificus, in the southwestern part of the East (Japan) Sea. Fish Res 91: 281-290.
-
8. Goldon AL, Giulivi CF. 2004. Pacific decadal oscillation and sea level in the Japan/East sea. Deep-Sea Res Part 1 51: 653-663.
-
9. Francis RC, Hare SR, Hollowed AB, Wooster WS. 1998. Effects of interdecadal climate variability on the oceanic ecosystems of the NE Pacific. Fish Oceanogr 7: 1-21.
-
10. Hare SR, Mantua NJ. 2000. Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog Oceanogr 47: 103-145.
-
11. Hong CH, Cho KD. 1983. The northern boundary of the Tsushima warm current and its fluctuations. J Oceanol Soc Kor 28: 1-9.
-
12. Jung HG. 2013. Regional differences in the response of ocean environment and fisheries resources in Korean waters to the North Pacific regime shifts and possible mechanisms, Master Dissertation, Gangneung -Wonju National University, Gangneung, Korea.
-
13. Kasahara S. 1978. Descriptions of offshore Squid Angling in the Sea of Japan, with Special Reference to the Distribution of Common Squid and on the Techniques for Forecasting Fishing Conditions. Bull Jap Sea Nat Fish Res Inst 29: 179-199.
-
14. Kawabata A, Yatsu A, Ueno Y, Suyama S, Kurita Y. 2006. Spatial distribution of the Japanese common squid, Todarodes pacificus, during its northward migration in the western North Pacific Ocean. Fish Oceanogr 15: 113-124.
-
15. Kidokoro H. 2009. Changes in the fishing grounds and fishing season of Japanese common squid Todarodes pacificus around Japanese waters. Rep Ann Meeting and Squid Res 3-10.
-
16. Kidokoro H, Goto T, Nagasawa T, Nishida H, Akamine T, Sakurai Y. 2010. Impact of a climate regime shift on the migration of Japanese common squid (Todarodes pacificus) in the Sea of Japan. ICES J Mar Sci 67: 1314-1322.
-
17. Kidokoro H, Sakurai Y. 2008. Effect of water temperature on gonadal development and emaciation of Japanese common squid Todarodes pacificus (Ommastrephidae). Fish Sci 74: 553-561.
-
18. Kim JJ. 2012. A study of early life stage for Japanese common squid (Todarodes pacificus) based on observations and modelling approaches. Ph. D. Dissertation, Pukyung National University, Busan, Korea: 1-73.
-
19. Kim YH, Moon CH, Choi KH, Lee CI. 2010. Relationship between squid (Todarodes pacificus) catch by sea block and marine environment in the East Sea during 1980s and 1990s. J Kor Soc Mar Env Safty 16: 259-268.
-
20. Kim YH, Moon CH, Lee CI. 2014. Distribution of the Common Squid Todarodes pacificus Paralavae in the Southern Coastal Waters in the East Sea in August and September 2013. J Kor Soc Oceanogr 19: 215-222.
-
21. Kim YH, Jung HK, Lee CI. 2018. Changes in the Spawning Ground Environment of the Common Squid, Todarodes pacificus due to Climate Change. Ocean and Polar Res 40: 127-143.
-
22. Kim YH, Choi KH, Lee CI. 2017. Migration and Distribution of the Common Squid (Todarodes pacificus) in Korean Waters. J Env Sci Int 26: 173-181.
-
23. Kim YH, Shin DH. 2019. Distribution of Common Squid Todarodes pacificus Larvae in the Southwestern Part of the East Sea in Summer and Autumn, 2015. J Kor Soc Mar Env Safty 25: 220 -228.
-
24. Kim YH, Lee CI. 2016. Distribution of water temperature and common squid Todarodes pacificus Paralavae around Korean Waters in 2013, 2014. J Kor Soc Mar Env Safety 22: 11-19.
-
25. Kim YH. 2015. The population ecology of the common squid (Todarodes pacificus) in the northwest Pacific Marginal Seas. Ph. D. Dissertation, Pukyung National University, Busan, Korea: 1-120.
-
26. Lee CI. 2003. Relationship between variation of the Tsushima Warm Current and current circulation in the East Sea. Ph.D. Thesis, Pukyung National University: 93.
-
27. Lee SD, Son YS, Kim YC. 1985. A Study on the vertical distribution of common squid, Todarodes pacificus (Steenstrup) in the eastern waters Korea. Bull Fish Res Agency 36: 23-28.
-
-
29. Mantua NJ, Hare SR, Zhang Y, Wallace JM, Francis RC. 1997. A Pacific Interdecadal Climate Oscillation with Impacts on Salmon Production. https://doi.org/10.1175/1520-0477(1997) 078<1069:APICOW>2.0.CO;2
-
30. Miller AJ, Chai F, Chiba S, Moisan JR, Neilson D. 2004. Decadal-scale climate and ecosystem interactions in the North Pacific Ocean. J Oceanogr 60: 163-188.
-
31. Minobe S. 1997. A 50-70 year climatic oscillation over the North Pacific and North America. Geophy Res Lett 24: 683-686.
-
32. Mokrin NM, Novikov YV, Zuenko YI. 2002. Seasonal migrations and oceanographic conditions for concentration of the Japanese flying squid (Todarodes pacificus STEENSTRUP, 1880) in the Northwesthern Japan Sea. Bull Mar Sci 71: 487-499.
-
-
34. Okutani T. 1983. Todarodes pacificus, in: Boyle, P. R. (ed.), Cephalopod Life Cycles, Academic, London, U. K., 1: 201-214.
-
35. Overland J, Rodionov S, Minobe S, Bond N. 2008. North Pacific regime shifts: Definitions, issues and recent transitions. Prog Oceanogr 77: 92-102.
-
36. Roper CFE, Sweeny MJ, Nauen CE. 1984. Family ommastrephidae, in: FAO species catalogue, 3, Cephalopods of the world, An annotated and illustrated catalogue of species of interest to fisheries, FAO Fisheries Synopsis: 227.
-
37. Pang IC, Oh KH. 2000. A Seasonal Circulation in the East China Sea and the Yellow Sea and its Possible Cause. J Kor Soc Oceanogr 35: 161-169.
-
38. Rosa AL, Yamamoto J, Sakurai Y. 2011. Effects of environmental variability on the spawning areas, catch, and recruitment of the Japanese common squid, Todarodes pacificus (Cephalopoda: Ommastrephidae), from the 1970s to the 2000s. ICES J Mar Sci 68: 1114-1121.
-
39. Sakurai Y, Kiyofuji H, Saitoh S, Goto T, Hiyama Y. 2000. Change in inferred spawning areas of Todarodes pacificus (Cephalopoa: Ommastrephidae) due to changing environmental conditions. ICES Journal of Marine Science 57: 24-30.
-
40. Sakurai Y. 2006. How climate change might impact squid populations and ecosystems: a case study of the Japanese common squid, Todarodes pacificus. Globec Report 24: 33-34.
-
41. Sakurai Y, Kiyofuji H, Saitoh S, Yamamoto J, Goto T, Mori K, Kinoshita T. 2002. Stock fluctuations of the Japanese common squid, Todarodes pacificus, related to recent climate changes. Fish Sci 68: 226-229.
-
42. Shevtsov GA, Zuev MA, Katugin ON, Mokrin NM. 2005. Distribution and biology of the Japanese common squid (Todarodes pacificus) in the Pacific Ocean off the Kuril Islands and eastern Kamchatka in summer. Ruthenica 15: 99-108.
-
43. Watanabe K, Sakurai Y, Segawa S, Okutani T. 1996. Development of an ommastrephid squid Todarodes pacificus from fertilized egg to the rhynchoteuthion paralarva. Amer Mal Bull 13: 73 88.
-
44. Yamamoto J, Shimura T, Uji R, Masuda S, Watanabe S, Sakurai Y. 2007. Vertical distribution of Todarodes pacificus (Cephalopoda Ommastrephidae) paralarvae near the Oki Islands, southwestern Sea of Japan. Mar Biol 153: 7-13.
Copyright (C) 2024. The Korean Society of Marine Life Science