










JMLS 2018 December;3(2):59-66. http://jmls.or.kr Epub 2018 December 17
Copyright © 2018 by The Korean Society of Marine Life Science
Evaluation of Health of Tidal Flat on the West Coast using Biomarker Genes and Manila Clam (Ruditapes philippinarum)
Hee Chan Choi; East Sea Fisheries Research Institute, NIFS, Gangneung 25435 Korea
Yoon Seok Choi; West Sea Fisheries Research Institute, NIFS, Incheon 22383 Korea
Han Seung Kang; Department of Genome Research, MS BioLab, Daejeon 34576, Korea
Yoon Lee; East Sea Fisheries Research Institute, NIFS, Gangneung 25435 Korea
- Abstract
갯벌의 건강 수준에 대한 평가는 갯벌에 서식하는 생물의 건강에 의해 평가될 수 있다. 생체지표유전자의 발현 분석을 통하여 갯벌에 서식하는 생물의 건강 수준을 평가할 수 있다. 본 연구의 목적은 heat shock protein 70 (Hsp70), heat shock protein 90 (Hsp90), glutathione S-transferases (GST) 및 thioredoxin (TRX)과 같은 생체지표유전자를 이용하여 서해안 갯벌의 건강을 평가하는 것이다. 이들 유전자는 스트레스, 면역 및 항산화 관련한 유전자들로서 이들 유전자의 발현을 통해 생물의 건강 정도를 관찰하는 데 사용할 수 있다. 본 연구에서는 서해안의 8개 정점에서 바지락(Ruditapes philippinarum)을 수집했다. 유전자의 발현은 RT-qPCR 방법으로 분석하였다. 연구 결과 Hsp70, Hsp90, GST 및 TRX 유전자들의 발현이 8개 정점에서 차별적으로 발현되는 것으로 나타났다. 특히, Hsp90 및 GST의 발현 또는 Hsp70 및 TRX의 발현은 유사하였다. 이것은 각 유전자에 특이적으로 반응하는 물질이 존재하는 것으로 생각된다. 따라서 이화학적 분석 결과에 근거하여 분석에 적합한 유전자를 선택할 수 있다고 생각한다. 이 결과는 Hsp70, Hsp90, GST 및 TRX 유전자는 갯벌의 건강을 평가하기 위한 생체지표유전자로서의 역할을 수행함을 시사한다.
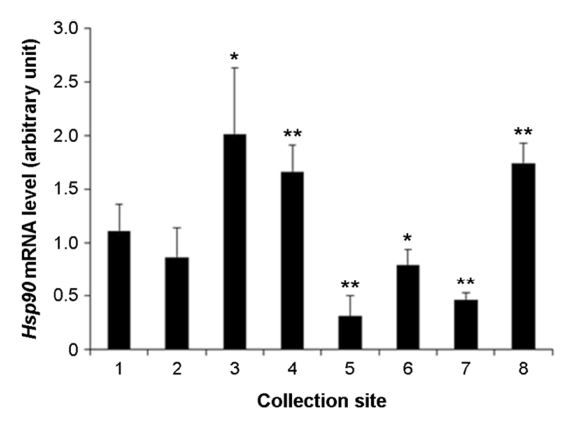
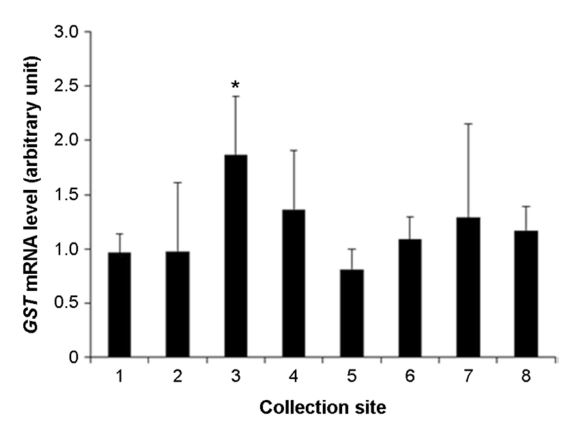
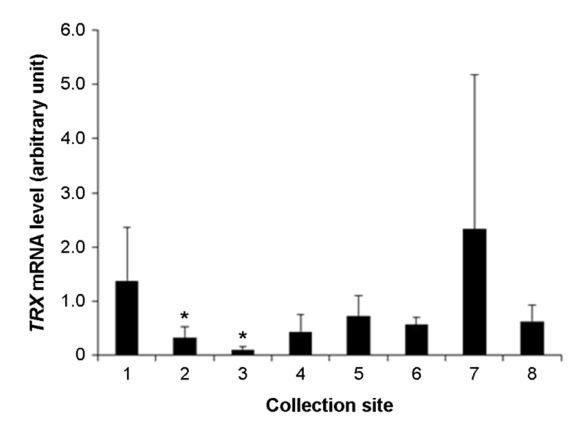
The assessment of level of health of the tidal flats can be evaluate by health of organisms inhabit the tidal flats. It is possible to evaluate the precise health level of organisms inhabit the tidal flats using analysis of expression of biomarker genes. The purpose of this research is to evaluate the health of the tidal flats on the west coast using biomarker genes such as heat shock protein 70 (Hsp70), heat shock protein 90 (Hsp90), glutathione S-transferases (GST) and thioredoxin (TRX). These genes are stress, immune, and antioxidant related genes that can be used to look at the health of an organism through gene expression. In this study, we collected manila clam (Ruditapes philippinarum) in 8 analysis areas on the west coast. Expression of the genes was analyzed by RT-qPCR method. Results showed that, the expression of Hsp70, Hsp90, GST and TRX genes were differentially expressed in the 8 analysis areas. In particular, the expression of Hsp90 and GST or the expression of Hsp70 and TRX were similar. This means that there is a substance that reacts specifically to each gene. Therefore, I think suggest that the based on the results of physicochemical analysis, it can be selected genes suitable for analysis. These results suggest that Hsp70, Hsp90, GST and TRX were played roles in biomarker for assessment of the health of tidal flats.
Keywords: Ruditapes philippinarum(바지락) Tidal flat(갯벌) Health(건강) RT-qPCR(역전사실시간장량적PCR) Biomarker gene(생체지표유전자)
Correspondence to: Yoon Lee; East Sea Fisheries Research Institute, NIFS, Gangneung 25435 Korea; E-mail : yoonlee@korea.kr;
- Received
- 10 December 2018;
- Revised
- 10 December 2018;
- Accepted
- 13 December 2018.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
갯벌의 어장 환경은 기후 변화 및 오염원 유입 등의 원인에 의해 과거와 비교하여 갯벌 어업의 생산성 하락 및 생물의 군집 변화 등이 나타나고 있다. 효과적이고 과학적인 갯벌 어장 환경을 관리하기 위하여 정부나 지방자치단체는 생물 서식 조사 및 이화학적 분석을 통한 조사 등의 모니터링을 시행하고 있다. 이러한 모니터링 조사 연구를 통해 환경기준설정의 기초자료 생산, 효율적인 사용방안 마련 및 복원 계획을 수립하고 있다. 자연생태계 환경모니터링 조사 방법의 일환으로 생체지표유전자(biomarker gene)의 활용이 부각되고 있다. 생물은 스트레스에 노출되면 항상성을 유지하기 위하여 세포 수준에서 스트레스 완화를 위한 기전이 작용하는데, 이에 관여하는 유전자를 개발 및 분석 평가하면 생물의 스트레스에 따른 건강 상태를 평가할 수 있다. 또한 생체지표유전자를 활용하여 평가하면 빠른 시간에 생물의 건강 상태를 평가할 수 있기에 저비용 및 고효율의 경제성도 장점이라 할 수 있다. 생체지표유전자로 활용 가능한 유전자의 개발은 다양한 기능의 유전자군에서 개발 가능하다. 해양생물에서는 수생태계 서식의 특성에 의해 수온, 염분 및 외래유입 오염물질에 따른 노출 스트레스에 대처한 항상성에 관여하는 유전자군으로서 열충격단백질(heat shock protein, Hsp), 산화스트레스(oxidative stress) 및 면역(immune) 관련 유전자군들이 주요하게 선정되고 있다.
Hsp는 온도 변화에 따른 세포 보호의 기전을 가진 단백질로 알려져 있으며 많은 연구가 이루어졌다. 최근에는 온도, 염분, 중금속(heavy metal), 내분비계장애물질(endocrine disrupting chemicals, EDCs) 및 산화스트레스(oxidative stress) 영향과의 연구 등을 통해 이들 인자와 Hsp 유전자 발현에 관한 연구 등이 보고되었다(Parsell and Lindquist, 1993; Feder and Hofmann, 1999; Kregel, 2002; Fangue et al., 2006; Colinet et al., 2010; Xu and Qin, 2012; You et al., 2013). Hsp는 면역 상태의 지표를 나타내는 단백질로서 분자량을 기준으로 Hsp110, Hsp100, Hsp90, Hsp70, Hsp60 및 저 분자량 Hsp 등으로 존재하고 있다(Li et al., 2009; Colinet et al., 2010; Xu and Qin, 2012; Teixeira et al., 2013). 이 중에서 Hsp70은 진화적으로도 잘 보존되어 있으며, 환경요인의 영향에 따라 발현에 차이를 보이는 대표적인 유전자이다(Chirico et al., 1988; Bukau and Horwich, 1998). 해양생물에서 환경요인 스트레스인 고수온과 빈산소 등은 Hsp70 유전자의 발현을 증가시킨다는 보고가 있다(Dong et al., 2008; Lee et al., 2012; Kim et al., 2015). Hsp90은 세포분화 및 발생에 관여하고, 스트레스에 반응 시 단백질조절, 세포사(apoptosis) 조절 및 신호전달에 관여한다고 알려져 있다(Jakob et al., 1995; Galea-Lauri et al., 1996; Imai and Yahara, 2000; Richter and Buchner, 2001; Soetaert et al., 2006; Wu and Chu, 2008; Li et al., 2009). 약물대사효소계 phaseⅡ로 알려진 glutathione S-transferase (GST)는 모든 생물체에 존재하며 다양한 기전을 가진 단백질이다. 온도(temperature), 염분(salinity) 등의 환경요인을 비롯하여 외인성생물화합물(xenobiotic compound), 중금속(heavy metal), 병원체(pathogen) 등에 의해 생성된 산화 스트레스(oxidative stress) 등의 독소를 체외로 배출시키거나, 반응성이 적은 물질로 분해하여 해독시키는 작용에 관여한다(Clark, 1989: Hayes and Pulford, 1995; Kampranis et al., 2000; Mueller et al., 2000; Agrawal et al., 2002; Smith et al., 2004; Hayes et al., 2005). GST의 이러한 작용기전은 독성시험의 지표로 활용되어 polychlorinated biphenyls (PCBs), dichloro-diphenyl-trichloroethane (DDT), polycyclic aromatic hydro- carbons (PAHs), 내분비계장애물질(endocrine disrupting chemicals; EDCs) 및 중금속 등과 GST와의 상관관계에 대한 연구가 많이 진행되었다(Looise et al., 1996: Hansson et al., 2006; Hamed et al., 2003; Koutsogiannaki et al., 2014; Luchmann et al., 2014; Zhang et al., 2014). 패류에서의 연구 결과로는 비단가리비(Chlamys farreri), 가리비(Chlamys islandica), 녹색홍합(Perna viridis), 굴(Crassostrea brasiliana), 지중해담치(Mytilus galloprovincialis) 등에서 환경요인 및 EDCs와 GST의 고발현에 관한 연구가 보고되었다(Lau and Wong, 2003; Luca-Abbott et al., 2005; Myrnes and Nilsen, 2007; Hu et al., 2015; Boukadida et al., 2017; Lima et al., 2018). Thioredoxin (TRX)은 원핵생물 및 진핵생물 모두에서 발견되는 단백질로서 활성산소종(reactive oxygen species, ROS)의 활성조절 효소로 면역학적으로 매우 중요하다고 알려져 있다(Liyanage et al., 2018). 활성산소는 산소의 정상적인 대사작용에 의해서 자연스럽게 생기며, 세포신호와 항상성에 중요한 역할을 하는데 환경적인 스트레스에 노출되면 매우 빠르게 증가하여 세포를 손상시키는데 이것이 산화 스트레스(oxidative stress)이다(Devasagayam et al., 2004). TRX는 산화 스트레스로 세포 손상이 발생하면 발현이 증가함으로써 산화 스트레스 상태에서 생체 방어 및 적응 반응을 하는데, 자체 환원력을 이용한 산화물을 제거하는 항산화 작용을 하거나 산화적 손상을 받은 단백질의 재생 및 산화-환원 반응을 통해 NF-kB (nuclear factor kappa B), AP-1 (activator protein-1) 전사인자 활성 조절, interleukin, tumor necrosis factor와 같은 cytokine의 생산을 증가시켜 세포증식 및 세포사(apoptosis)를 방지하는 등 여러 생물학적 현상을 유발함으로써 산화-환원 반응에 의한 생체의 신호전달체계에서 중심적인 역할을 하는 단백질이다(Spector et al., 1988; Fernado et al., 1992; Mitsui et al, 1992; Yodoi and Uhyama, 1992; Schenk et al., 1994).
본 연구의 목적은 갯벌생물 중에서 서해안에 많이 서식하는 양식생물인 바지락(Ruditapes philippinarum)을 대상으로 갯벌어장의 환경에 의해 발생 가능한 스트레스로 인한 건강 정도를 생체지표유전자를 이용하여 바지락 개체의 건강 상태를 평가함과 동시에 바지락이 서식하는 갯벌의 건강 도를 평가하고자 한다. 평가에는 Hsp70, Hsp90, GST 및 TRX를 생체지표유전자로 선정하였고 이들 유전자의 상대적 발현을 통해 평가를 하였다.
재료 및 방법
1. 실험동물 및 시료
서해안 갯벌어장 8개 정점(중앙, 주교, 원청, 만풍, 선유도, 백미리, 파도리, 고남리)에서 2018년 6월 바지락을 채집하였다. 채집한 바지락은 염수에 불순물을 세척한 후, 패각을 열어 내장을 포함한 육질 부위를 분리 수집하였다. 수집한 바지락 육질 부위는 생리식염수를 이용하여 세척한 후, 멸균 필터페이퍼를 이용하여 수분을 제거하였다. 수분이 제거된 육질 부위는 RNAlater (Life technologies, Carlsbad, CA, USA) 용액이 첨가된 용기에 담은 후, 저온에서 운반 및 4℃ 보관한 후 실험에 사용하였다. 시료 준비를 위한 바지락은 정점당 10개체를 채집하여 사용하였다.
2. Total RNA 추출
Total RNA 추출은 RNAiso Plus (TaKaRa Co. Shiga, Japan) 시약 용액을 사용하였다. 저온 보존된 바지락 조직을 액체질소에 침지시킨 후, 막자사발을 이용해 마쇄시켰다. 마쇄된 조직은 RNAiso plus 용액(1 ml/100 mg)이 들어있는 tube에 넣고 혼합한 후 실온에서 5분간 방치하였다. Chloroform 0.2 ml를 넣어주고 혼합 후 실온에서 2분간 방치하였다. 원심분리를 시행하여 상등액을 새로운 tube에 옮겨 담고 isopropyl alcohol 1.0 ml를 넣어 10분간 실온에서 방치하였다. 원심분리를 시행하여 pellet를 생성시킨 후, 상등액을 제거하고 75% EtOH로 세척한 후 DEPC-DW로 녹여 추출하였다. 추출한 total RNA는 분광광도계(NanoVue, GE Healthcare, Boston, MA, USA)를 이용하여 정성 및 정량하였고, RNA quality는 260/280 ratio 1.8 이상을 확인하였으며, 역전사 반응(Reverse Transcription: RT) 전까지 -80℃ 초저온 냉동고에 보관하였다. 정점당 채집한 10개체에서 추출한 total RNA는 동량을 얻은 후, 합쳐서 실험에 사용하였다.
3. 중합효소연쇄반응(RT-qPCR)
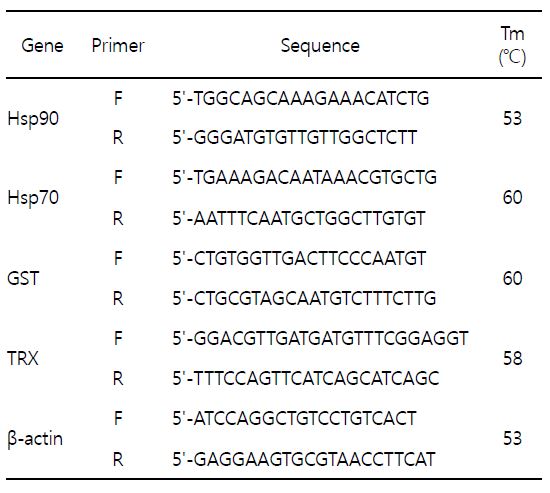
cDNA 합성을 위한 역전사 반응(Reverse Transcription, RT)은 iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA)를 이용하였다. Total RNA 1 μg, iScript 5×Master mix 4 μl, iScript reverse transcriptase 1 μl 및 DEPC-DW를 넣어 최종 반응 용액 20 μl을 맞춘 후, 42℃에서 1시간 반응하여 cDNA를 합성하였다. 중합효소연쇄반응(qPCR)은 iQ SYBR Green Supermix kit (Bio-Rad, Hercules, CA, USA)를 이용하여 수행하였다. cDNA 1 μl, primer 각각 1 μl, iQ SYBR Green Supermix (2×) 10 μl 및 DEPC-DW를 넣어 최종 반응 용액 20 μl 되게 맞춘 후, real-time PCR machine (CFX96, Bio-Rad, Hercules, CA, USA)를 이용하여 증폭하였다. 생체지표유전자로 선정된 바지락 특이적인 유전자인 Hsp70 (Genbank Accession No: KM 280636), Hsp90 (Genbank Accession No: KJ 569080), GST (Genbank Accession No: JN 593116) 및 TRX (Genbank Accession No: JF 499393)의 염기서열을 바탕으로 Primer3Plus 프로그램을 이용하여 프라이머를 제작하였다. PCR 반응을 위한 생체지표유전자 프라이머의 염기서열 및 annealing temperature는 Table 1에 명기하였다. 유전자 증폭을 위한 반응 조건은 95℃에서 5분 1회, 95℃에서 30초, 각 유전자 annealing temperature에서 30초, 72℃에서 30초를 35회 반복하였으며, 마지막으로 72℃에서 10분간 1회 반응하였다. Melting curve의 분석은 0.5℃ 간격으로 50℃에서부터 95℃까지 상승시켰다가, 이후 30℃에서 5분간 유지하였다. 상대적인 유전자 발현량의 결정은 2-∆∆Ct 방법(comparative Ct method)을 이용하여 유전자의 발현량을 분석하였다. 내재표준유전자로는 house keeping 유전자인 β-actin (ACTB)을 사용하여 발현량을 normalization시켰다. 실시간 중합효소연쇄반응은 5회 반복 실험하였다.
4. 통계학적 분석
실험군간의 유의성 검정은 Student's t-test로 비교하였으며, p가 0.05 이하인 것만 유의한 것으로 하였다.

- References
-
1. Agrawal GK, Jwa NS, Rakwal R. 2002. A pathogen-induced novel rice (Oryza sativa L.) gene encodes a putative protein homologous to type II glutathione S-transferases. Plant Sci 163: 1153-1160.


-
2. Ahlf W, Holler H, Neumann-Hense H, Ricking M. 2002. A guidance for the assessment and evaluation of sediment quality: A german approach based on ecotoxicological and chemical measurements. J Soils Sediment 2: 37-42.
-
3. Atici T, Ahiska S, Altindag A, Aydin D. 2008. Ecological effect of some heavy metals (Cd, Pb, Hg, Cr) pollution of phytoplanktonic algae and zooplanktonic organisms in Sariyar Dam Reservoir in turkey. Afr J Biotechnol 7: 1972-1977.
-
4. Beiras R, femández N, Bellas J, Besada V, González-Quijano A, Nunes T. 2003. Inttegrative assessment of marine pollution in Galician estuaries using sediment chemistry, mussel bioaccumulation, and embryo-larval toxicity bioassays. Chemosphere 52: 1209-1224.
-
5. Boukadida K, Cachot J, Clérandeaux C, Gourves PY, Banni M. 2017. Early and efficient induction of antioxidant defense system in Mytilus galloprovincialis embryos exposed to metals and heat stress. Ecotoxicol Environ Saf 138: 105-112.
-
-
7. Chirico WJ, Waters MG, Blobel G. 1988. 70 K heat shock related proteins stimulate protein translocation into microsomes. Nature 332: 805-810.
-
8. Chu KW, Chow KL. 2002. Synergistic toxicity of multiple heavy metal is revealed by a biological assay using a nematode and its transgenic derivative. Aquat Toxicol 61: 53-64.
-
9. Clark AG. 1989. The comparative enzymology of the glutathione S-transferases from non-vertebrate organisms. Comp Biochem Physiol 92: 419-446.
-
10. Colinet H, Lee SF, Hoffmann A. 2010. Temporal expression of heat shock genes during cold stress and recovery from chill coma in adult Drosophila melanogaster. FEBS J 277: 174-185.
-
11. DeForest DK, Brix KV, Adams WJ. 2007. Assessing metal bioaccumulation in aquatic environments: the inverse relationship between bioaccumulation factors, trophic transfer factors and exposure concentration. Aquat Toxicol 84: 236-246.
-
12. Devasagayam TP, Tilak JC, Boloor KK, Sane KS, Ghaskadbi SS, Lele RD. 2004. Free radicals and antioxidants in human health: current status and future prospects. J Assoc Physicians India 52: 794-804.
-
13. Dong YW, Dong SL, Ji TT. 2008. Effects of different thermal regimes on growth and physiological performance of the sea cucumber Apostichopus japonicus Selenka. Aquaculture 275: 329 -334.
-
14. Fangue NA, Hofmeister M, Schulte PM. 2006. Intraspecific variation in thermal tolerance and heat shock protein gene expression in common killifish, Fundulus heteroclitus. J Exp Biol 209: 2859 -2872.
-
15. Feder ME, Hofmann GE. 1999. Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Annu Rev Physiol 61: 243-282.
-
16. Fernando MR, Nanri H, Yoshitake S, Nagata-Kuno K, Minakami S. 1992. Thioredoxin regenerates proteins inactivated by oxidative stress in endothelial cells. Eur J Biochem 207: 917-922.
-
17. Galea-Lauri J, Richardson AJ, Latchman DS, Katz DR. 1996. Increased heat shock protein 90 (hsp90) expression leads to increased apoptosis in the monoblastoid cell line U937 fol- lowing induction with TNF-alpha and cycloheximide: a pos- sible role in immunopathology. J Immunol 157: 4109-4118.
-
18. Hamed RR, Farid NM, Elowa SE, Abdalla AM. 2003. Glutathione related enzyme levels of freshwater fish as bioindicators of pollution. The Environmentalist 23: 313-322.
-
19. Han TJ, Han YS, Park GS, Lee SM. 2008. Development marine ecotoxicological standard methods for Ulva sporulation test. Kor J Soc Ocean 13: 121-128.
-
20. Hansson T, Schiedek D, Lehtonen KK, Vuorinen P, Liewenborg J, Noaksson E, Tjarnlund U, Hansson M, Balk L. 2006. Biochemical biomarkers in adult female perch (Perca fluviatilis) in a chronically polluted gradient in the Stockholm recipient (Sweden). Marine Pollution Bulletin 53: 451-468.
-
21. Hayes JD, Pulford DJ. 1995. The glutathione S-transferase supergene family: regulation of GST and the contribution of the isoenzymes to cancer chemprotection and drug resistance. Critical Reviews in Biochemistry and Molecular Biology 30: 445-600.
-
22. Hayes JD, Flanagan JU, Jowsey IR. 2005. Glutathione transferases. Annual Review of Pharmacology and Toxicology 45: 51-88.
-
23. Hu F, Pan L, Xiu M, Jin Q, Wang G, Wang C. 2015. Bioaccumulation and detoxification responses in the scallop Chlamys farreri exposed to tetrabromobisphenol A (TBBPA). Environ Toxicol Pharmacol 39: 997-1007.
-
24. Imai J, Yahara I. 2000. Role of HSP90 in salt stress tolerance via stabilization and regulation of calcineurin. Mol Cell Biol 20: 9262-9270.
-
25. Jakob U, Lilie H, Meyer I, Buchner J. 1995. Transient interaction of Hsp90 with early unfolding intermediates of citrate synthase. J Biol Chem 270: 7288-7294.
-
26. Kampranis SC, Damianova R, Atallah M, Toby G, Kondi G, Tsichlis PN, Makris AM. 2000. A novel plant glutathione S-transferase /peroxidase suppresses Bax lethality in yeast. J Biol Chem 275: 29207-29216.
-
27. Kim CW, Jin YG, Kim TI, Jeong DS, Kang HS. 2015. The expression of Hsp90 and ferritin genes under thermal stress in the sea cucumber (Apostichopus japonicas). Korean J Environ Biol 33: 433-440.
-
28. Koutsogiannaki S, Franzellitti S, Fabbri E, Kaloyianni M. 2014. Oxidative stress parameters induced by exposure to either cadmium or 17 beta estradiol on Mytilus galloprovincialis hemocytes. The role of signaling molecules. Aquat Toxicol 146: 186-195.
-
29. Kregel KC. 2002. Heat shock proteins: modifying factors in physiological stress responses and acquired thermotolerance. J Appl Physiol 92: 2177-2186.
-
30. Lau PS, Wong HL. 2003. Effect of size, tissue and location on six biochemical markers in the green lipped mussel, Perna viridis. Mar Pollut Bull 46: 1563-1572.
-
31. Lee G, Hwang J, Chung Y, Kim D, Moh SH, Chang M, Lee TK. 2012. Effects of hypoxia on the fertilization and early development of sea urchin, Strongylocentrous nudus. Journal of the Korea Academia-Industrial Cooperation Society 13: 3785-3791.
-
32. Li P, Zha J, Zhou KY. 2009. Molecular cloning, mRNA expression, and characterization of HSP90 gene from Chinese mitten crab Eriocheir japonica sinensis. Comp Biochem Physiol B 153: 229 -235.
-
33. Lima D, Zacchi FL, Mattos JJ, Flores-Nunes F, Gomes CHAM, de Mello ÁCP, Siebert MN, Piazza CE, Taniguchi S, Sasaki ST, Bícego MC, Bebianno MJ, de Almeida EA, Bainy ACD. 2018. Molecular and cellular effects of temperature in oysters Crassostrea brasiliana exposed to phenanthrene. Chemosphere 209: 307-318.
-
34. Lin H, Sun T, Zhou Y, Gu R, Zhang X, Yang W. 2018. Which genes in a typical intertidal seagrass (Zostera japonica) indicate copper-, lead-, and cadmium pollution? Front Plant Sci 9: 1545.
-
35. Liyanage DS, Omeka WKM, Godahewa GI, Lee J. 2018. Molecular characterization of thioredoxin-like protein 1(TXNL1) from big-belly seahorse Hippocampus abdominalis in response to immune stimulation. Fish Shellfish Immunol 75: 181-189.
-
36. Looise Bas AS, Holwerda DA, Foekema EM. 1996. Induction of glutathione S-transferase in the freshwater bivalve Sphaerium corneum as biomarker for short-term toxicity tests? Comp Biochem Physiol C 113: 103-107.
-
37. Luca-Abbott SB, Richardson BJ, McClellan KE, Zheng GJ, Martin M, Lam PKS. 2005. Field validation of antioxidant enzyme biomarkers in mussels (Perna viridis) and clams (Ruditapes philippinarum) transplanted in Hong Kong coastal waters. Mar Pollut Bull 51: 694-707.
-
38. Luchmann K, Dafre AL, Trevisan R, Craft JA, Meng X, Mattos JJ, Zacchi FL, Dorrington TS, Schroeder DC, Bainy ACD. 2014. A light in the darkness: New biotransformation genes, antioxidant parameters and tissue-specific responses in oysters exposed to phenanthrene. Aquat Toxicol 152: 324-334.
-
39. Lv J, Wang Y. 2018. Multi-scale analysis of heavy metals sources in soils of Jiangsu Coast, Eastern China. Chemosphere 212: 964-973.
-
40. Meng X, Tian X, Liu M, Nie G, Jiang K, Wang B, Wang L. 2014. The transcriptomic response to copper exposure by the gill tissue of Japanese scallops (Mizuhopecten yessoensis) using deep-sequencing technology. Fish Shellfish Immunol 38: 287-293.
-
41. Meng X, Tian X, Nie G, Wang J, Liu M, Jiang K, Wang B, Guo Q, Huang J, Wang L. 2015. The transcriptomic response to copper exposure in the digestive gland of Japanese scallops (Mizuhopecten yessoensis). Fish Shellfish Immunol 46: 161 -167.
-
42. Mitsui A, Hirakawa T, Yodoi J. 1992. Reactive oxygen reducing and protein-refolding activities of adult T cell leukemia-derived factor/human thioredoxin. Biochem Biophy Re Sear Comm 186: 1220-1226.
-
43. Mueller LA, Goodman CD, Silady RA, Walbot V. 2000. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. Plant Physiol 123: 1561-1570.
-
44. Myrnes B, Nilsen IW. 2007. Glutathione S-transferase from the Icelandic scallop (Chlamys islandica): isolation and partial characterization. Comp Biochem Physiol C Toxicol Pharmacol 144: 403-407.
-
45. Parsell D, Lindquist S. 1993. The function of heat-shock proteins in stress tolerance: degradation and reactivation of damaged proteins. Annu Rev Genet 27: 437-496.
-
46. Richter K, Buchner J. 2001. Hsp90: chaperoning signal transduction. J Cell Physiol 188: 281-290.
-
47. Schenk H, Klein M, Erdbrugger W, Droge W, Schulze-Osthoff K. 1994. Distinct effects of thioredoxin and antioxidants on the activation of transcription factors NF- kB and AP-1. Proc Natl Acad Sci USA 91: 1672-1676.
-
48. Sivula L, Vehniäinen ER, Karjalainen AK, Kukkonen JVK. 2018. Toxicity of biomining effluents to Daphnia magna: Acute toxicity and transcriptomic biomarkers. Chemosphere 210: 304-311.
-
49. Smith AP, De Ridder BP, Guo WJ, Seeley EH, Regnier FE, Goldsbrough PB. 2004. Proteomic analysis of Arabidopsis glutathione S-transferases from benoxacor- and copper- treated seedlings. J Biol Chem 279: 26098-26104.
-
50. Soetaert A, Moens LN, Van der Ven K, Van Leemput K, Naudts B, Blust R, De Coen WM. 2006. Molecular impact of propiconazole on Daphnia magna using a reproduction-related cDNA array. Comp Biochem Physiol C 142: 66-76.
-
51. Spector A, Yan GZ, Huang RRC, Macdermott MJ, Gascoyne PRC, Pigiet V. 1988. The effect of H2O2 upon thioredoxin-enriched lens epithelial cells. J Biol Chem 263: 4984-4990.
-
52. Taylor DA, Nair SV, Thompson EL, Raftos DA. 2015. Dosedependent effects of metals on gene expression in the sydney rock oyster, Saccostrea glomerata. Environ Toxicol 30: 989-998.
-
53. Teixeira T, Diniz M, Calado R, Rosa R. 2013. Coral physiological adaptations to air exposure: Heat shock and oxidative stress responses in Veretillum cynomorium. Journal of Experimental Marine Biology and Ecology 439: 35-41.
-
54. Wu LT, Chu KH. 2008. Characterization of heat shock protein 90 in the shrimp Metapenaeus ensis: evidence for its role in the regulation of vitellogenin synthesis. Mol Reprod Dev 75: 952 -959.
-
55. Xu Q, Qin Y. 2012. Molecular cloning of heat shock protein 60 (PtHSP60) from Portunus trituberculatus and its expression response to salinity stress. Cell Stress Chaperones 17: 589 -601.
-
56. Yodoi J, Uhiyama T. 1992. Diseases associated with HTLV-I virus, IL-2 receptor dysregulation and redox regulation. Immunol Today 13: 405-411.
-
57. You L, Ning X, Liu F, Zhao J, Wang Q, Wu H. 2013. The response profiles of HSPA12A and TCTP from Mytilus galloprovincialis to pathogen and cadmium challenge. Fish Shellfish Immunol 35: 343-350.
-
58. Zhang H, Pan L, Tao Y. 2014. Toxicity assessment of environmental pollutant phenanthrene in clam Venerupis philippinarum using oxidative stress biomarkers. Environ Toxicol Pharmacol 37: 697-704.
-
59. Zhang M, Bao ZY, Chen GG, Yong TJ, Zhu YP, Liang XH. 2017. Characteristics and risks of heavy metals content in surface sediment of tidal flat areas in eastern china. Huan Jing Ke Xue 38: 4513-4524.
Copyright (C) 2024. The Korean Society of Marine Life Science