










JMLS 2017 April;2(1):12-19. Epub 2017 May 11
Copyright © 2017 by The Korean Society of Marine Life Science
수온별 넙치(Paralichthys olivaceus)의 스트레스 반응 및 산소소비 변화(Change on Stress Responses and Oxygen Consumption of Olive Flounder, Paralichthys olivaceus at Different Water Temperature)
박형준; 국립수산과학원 양식관리과
김성연; 국립수산과학원 양식관리과
양성진; 국립수산과학원 양식관리과
민병화; 국립수산과학원 양식관리과
- Abstract
급격한 수온의 변화는 어류의 생리학적인 측면에서 스트레스를 유발한다. 본 연구에서는 넙치(Paralichthys olivaceus)로부터 각 수온별(9, 12, 15, 18 및 21℃) 조건에 따라 24 및 48시간 동안 노출시킨 후에, 혈액생리학적 분석, 스트레스 단백질로 알려진 Hsp70 mRNA 발현 및 산소소비량을 조사하였다. 혈액학적 분석에서 hematocrit (Ht) 및 hemoglobin (Hb), 혈장 코티졸 및 글루코스의 변화, aspartate aminotransferase (AST) 및 alanine aminotransferase (ALT), NH3, 삼투질농도(osmolality) 및 총단백질(total protein, TP)은 9℃ 및 12℃에서 다른 수온별 실험구에 비해 대부분의 항목에서 유의적인 차이를 보였다. Hsp70 mRNA 발현은 9℃ 및 12℃에서 다른 실험구에 비해 높은 발현량을 확인하였고, 산소소비량은 9℃ 및 12℃에서 21℃에 비해 낮았다. 이러한 결과는 넙치 종자의 장거리 수송을 위한 수온자료로 활용할 수 있다.
This study investigated physiological and hematological changes, expression of stress protein Hsp70 mRNA and oxygen consumption in olive flounder (Paralichthys olivaceus) after exposing the fish at different temperature conditions (9, 12, 15, 18 and 21℃) for 24 and 48 hours. Hematological parameters including hematocrit (Ht) and hemoglobin (Hb), cortisol and glucose, aspartate aminotransferase (AST) and alanine aminotransferase (ALT), NH3, osmolality and total protein (TP) mostly exhibited significant changes at 9 and 12℃. The expression of Hsp70 mRNA was also higher at 9 and 12℃ than at other temperatures. The measured oxygen consumptions were also lower at 9 and 12℃ than at 21℃. It is expected that the study results could be utilized as baseline data to control water temperature during long-distance transportation, e.g. for exporting olive flounder juveniles to overseas
Keywords: Olive flounder Paralichthys olivaceus(넙치) Blood physiological response(혈액생리학적 반응) Hsp70 Oxygen consumption(산소소비량)
Correspondence to: Byung-Hwa Min; Aquaculture Research Division, National Institute of Fisheries Science, Busan 46083 Korea; E-mail : pkmbh@korea.kr;
- Received
- 7 April 2017;
- Revised
- 10 April 2017;
- Accepted
- 20 April 2017.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Language: Korean/English,
Full Text:

서 론
환경적인 요인에 있어 수온의 변화는 어류에게 스트레스를 유발하고, 생리학적인 활성에 영향을 미치며, 지나친 수온의 자극은 폐사를 유발하기도 한다(Cossins et al., 1995; Ellis, 2001; Bowden, 2008). 수온의 변화로 인해 1차적으로 스트레스가 발생하게 되면 시상하부 및 뇌하수체의 활성이 증가함에 따라, 혈액 내 cortisol의 분비가 증가하게 된다(Wedemeyer and Mcleay, 1981; Perry and Reid, 1993; Chang and Hur, 1999). 2차적으로는 간세포 파괴에 의한 AST (aspartate aminotransferase) 및 ALT (alanine aminotrans- ferase), glucose와 NH3의 증가가 경골어류로부터 보고되어진 바 있다(Park et al., 2016; Yang et al., 2017).
또한 어류에게 수온변화로 인한 스트레스가 발생됨에 따라, 스트레스 관련 단백질로 알려진 heat shock proteins (Hsps)의 발현이 증가한다는 보고가 있다(Nolan et al., 1999; Choi et al., 2007). Hsp70은 모든 생물체에서 계통발생학적으로 잘 보존되어지고 있는 단백질이며, 주로 환경적인 변화(저산소, 수온 및 염분)와 미생물의 감염으로 인해 발현이 유도된다(Beckmann et al., 1990; Miliani et al., 2002; Srivastava, 2002). 이러한 스트레스 요인의 발생으로 인한 어류의 생리 및 생태학적 조건에 영향을 미치며, 세포 내 Hsps의 합성과 함께 다양한 생리학적 변화가 발생한다(Schlesinger et al., 1992). Hsp70은 스트레스 요인에 의한 충격으로부터 생체를 보호하기 위한 항상성(homeostasis) 유지에 중요한 역할을 수행한다(Forsyth et al., 1997; Iwama et al., 1998).
또한 생물이 소비하는 용존산소(dissolved oxygen)의 양은 생체 내 신진대사와 밀접한 관련이 있으며, 대사량의 변화를 알아보기 위한 척도로 활용되고 있다(Yang, 2017). 따라서, 안전한 활수산물의 수출 및 장거리 수송을 위해서는 대사량을 감소시켜야 함으로 수온별 산소소비량의 조사는 반드시 필요하다. 이전의 연구에 따르면 고수온 및 저수온에 따른 어류의 산소소비량 변화가 보고되어진 바 있지만(Do et al., 2016; Yang, 2017), 다양한 환경변화와 장거리 수송 시 적정환경을 구명하는데 있어 아직까지는 연구가 미흡한 실정이므로, 추후 계속적인 연구가 이루어져야 한다.
본 연구에서는 한국의 주요 해산 양식어종인 넙치로부터 수온변화에 따른 혈액학적 반응, 산소소비량의 변화 및 Hsp70 mRNA 발현패턴을 분석함으로써 넙치의 장거리 수송기술에 있어 적정수온을 구명하는데 기초적인 자료로 제시하고자 한다.
재료 및 방법
1. 실험어 및 실험조건
넙치(22.6±1.2 cm, 124.3±24.4 g)는 국립수산과학원의 생물 사육실에서 2주 동안 유수식 1 ton 원형수조에서 상업용 사료를 급이하며 순치시켰다. 수온은 21℃를 유지하였다. 이 후, 실험어 10마리를 가온 및 냉각이 가능한 50 ℓ 사각수조에 각각 수용한 뒤, 21℃에서 1시간마다 3℃씩 낮추어 각 수온별(9, 12, 15, 18 및 21℃)로 24 및 48시간 동안 노출시켰다. 모든 실험은 2반복으로 수행하였다.
2. 혈액분석
24 및 48시간 노출 후, 모든 실험어는 150 mg/l의 tricaine methan sulphonate (MS-222, Sigma, USA)로 마취시킨 다음, heparin sodium (Sigma, USA) 처리된 1 ml syringe를 이용하여 실험어의 미부정맥으로부터 혈액을 샘플하였다. 샘플 후, 혈액의 일부를 hematocrit (Ht) 및 hemoglobin (Hb) 분석에 사용하고 나머지 혈액은 실온에서 20분간 방치한 다음 원심분리(4℃, 10,000 rpm, 15분)하여 분석 전까지 deep-freezer (-80℃)에 보관하였다. Ht는 모세관을 원심분리(RT, 10,000 rpm, 15분)하여 Ht-log chip (Micro-haematocrit reader, Hawksley Co, UK)을 이용하여 측정하고, Hb, 혈장 글루코스, 총 단백질(TP), aspartate aminotransferase (AST), alanine aminotransferase (ALT) 및 NH3는 자동 생화학 분석기(Fuji dry-chem 4000i, Fujifilm Co., Japan)로 분석하였다. 삼투질농도는 삼투압 측정기(Vapro 5520, WESCOR Co., USA)로 측정하였다. 혈장 코티졸 농도는 Fish ELISA Cortisol kit (Cusabio Biotech, China)의 상법에 따라 분석하였다.
3. Quantitative real-time PCR (q-PCR)
24 및 48시간 동안 수온별로 노출시킨 넙치의 Hsp70 mRNA 발현을 분석하기 위해 각 실험구의 넙치로부터 간 조직을 적출하여 액체질소에서 급속 동결시킨 후, -80℃에서 실험 전까지 보관하였다. Total RNA는 TRizol Reagent (Gibco/BRL, USA)법을 이용하여, 각 시료에서 RNA를 추출한 후, 1 μg의 total RNA는 Transcriptor First Strand cDNA synthesis kit (Roche, USA)를 이용하여 cDNA를 합성하였다.
PCR 분석에 앞서, 이전에 알려진 넙치 Hsp70 nucleotide sequence (accession no. AAC33859)에 기초하여 primer를 제작하였다(Table 1). Hsp70 mRNA 발현은 CFX96TM Real-time System (Bio-Rad, USA)과 iQTMSYBR green Supermix (Bio-Rad, USA)를 이용하여 분석하였다. 대조구로써 house-keeping gene은 넙치에서 알려진 β-actin (accession no. HQ386788)을 사용하였으며, 모든 분석값은 β-actin에 대한 cycle (Ct) level과의 차이로 계산하였다. Ct 값은 다음과 같은 식으로 결정지었다: 2-ΔΔCt Method, [ΔΔCt=2^-(ΔCtsample - ΔCtinternalcontrol)] (Livak and Schmittgen, 2001).

4. 산소소비량 측정
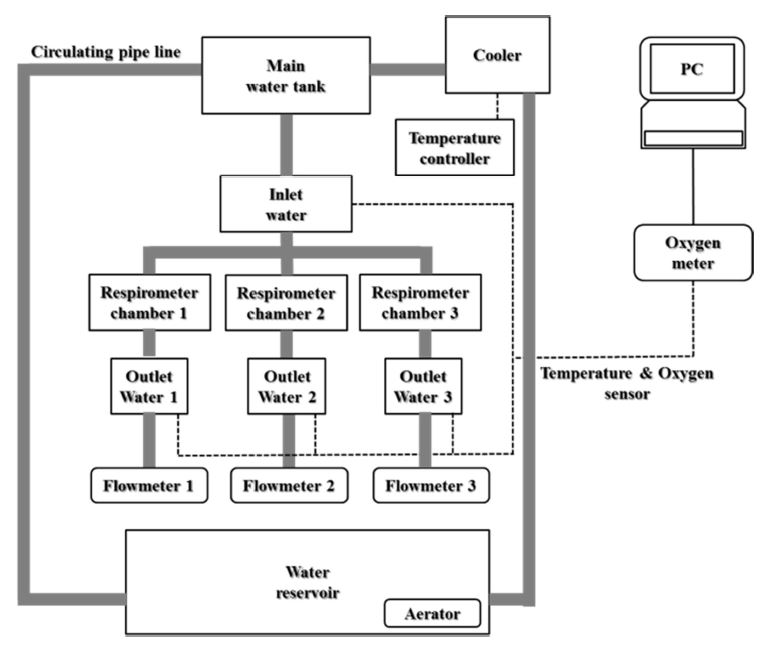
수온별로 노출시킨 넙치의 호흡에 따른 산소소비량을 조사하기 위해 호흡측정챔버(respirometry)를 사용하였고, 유입수와 유출수 간 용존산소의 농도차이를 이용하여 측정값을 구하였다. 호흡측정챔버는 가로 25 cm, 세로 30 cm 및 높이 17 cm로 부피가 약 12.8 ℓ인 직육면체의 아크릴 수조로 제작되었다. 실험에 사용될 넙치는 48시간 동안 절식 후, 호흡측정챔버에 넣었으며, 각 수온별 용존산소량은 생물호흡측정기(Oxygen Optode Sensor 3835, Aandreaa Data Instrument Inc, Norway)를 이용하였다. 10분 간격으로 24 및 48시간 동안 생물호흡측정기와 연결된 컴퓨터를 통해 Oxygen monitoring software program을 이용하여 측정된 수치를 바탕으로 산소소비량의 평균값을 구하였다. 그 식은 다음과 같으며, 호흡측정시스템은 Fig. 1에 나타내었다.
• 단위체중당 산소소비량(mg O2/kg/hr) ={(Ci-Co)×F×60}/W
• Ci = 유입수의 용존산소량(mg/l)
• Co = 유출수의 용존산소량(mg/l)
• F = 유량(L/min)
• W = 체중(kg)
5. 통계분석
결과값은 평균 표준편차±(S.E.)로 나타내었으며, 실험구별 유의차 유무는 SPSS 통계처리프로그램(version 10.0; SPSS inc., USA)에 의한 One-way ANOVA 및 Duncan's multiple range test를 실시하여 평균간의 유의성(p < 0.05)을 검정하였다.
결 과
1. 혈액생리학적 분석
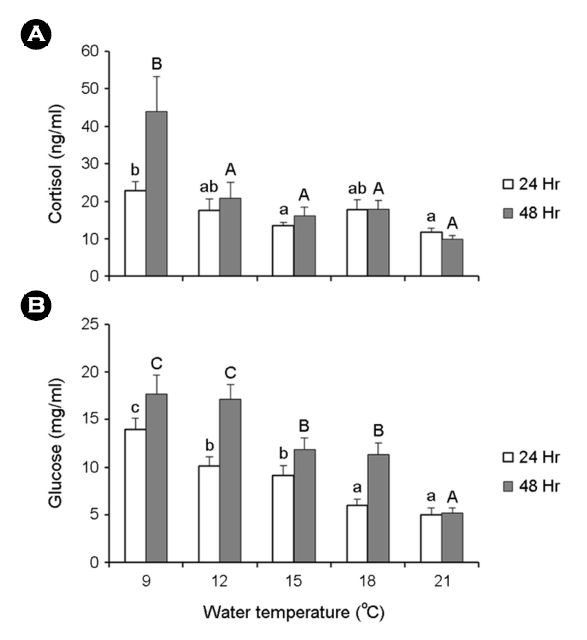
수온별로 노출시킨 넙치의 혈액생리학적 반응을 확인하였다. 코티졸 농도는 24 및 48시간 노출 후, 9℃에서 각각 22.8±2.5, 43.9±9.3 ng/ml로 대조구인 21℃보다 유의하게 높은 값을 나타내었고, 글루코스의 변화도 마찬가지로 22.8±2.5, 43.9±9.3 ng/ml로 다른 실험구에 비해 유의한 차이를 나타내었다(Fig. 2).
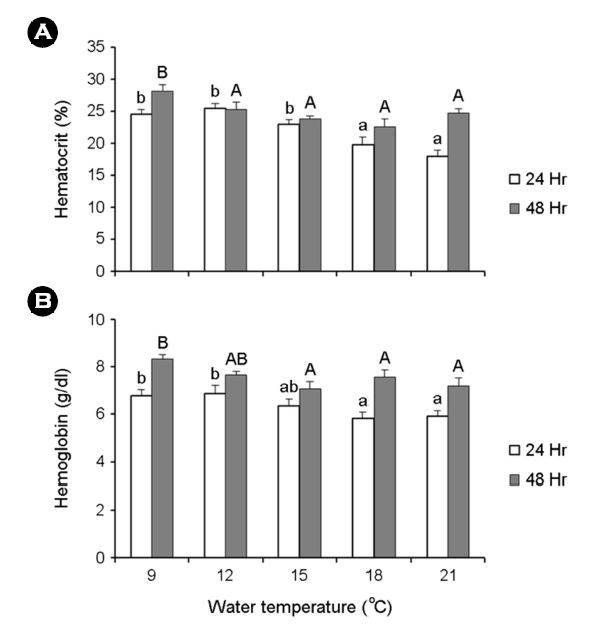
Ht는 24시간 노출 후, 12℃에서 25.4±0.7%, 48시간 노출 후, 9℃에서 28.2±0.9%로 다른 실험구에 비해 유의하게 차이를 보였다. 또한 Hb에서도 24시간 노출 후, 12℃에서 6.9±0.4 g/dl, 48시간 노출 후, 9℃에서 8.3±0.2 g/dl로 대조구인 21℃에 비해 유의한 차이를 보였다(Fig. 3).
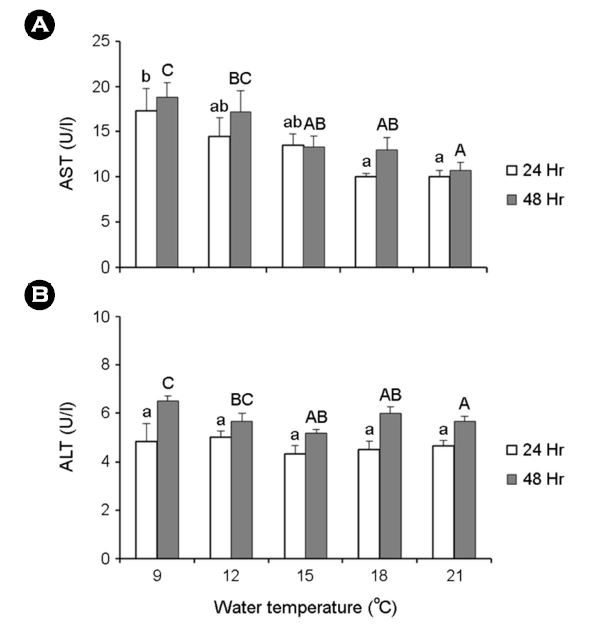
24 및 48시간 노출 후의 AST는 9℃에서 각각 17.3±2.5 및 18.8±1.6 U/l로 나타났고, ALT는 24시간 노출 후에는 실험구 간의 유의한 차이를 보이지 않았지만, 48시간 노출 후의 9℃에서 6.5±0.2 U/l로 대조구인 21℃에 비해 유의하게 증가하였다(Fig. 4).
총단백질, NH3, 삼투질농도, Na+, K+ 및 Cl-은 Table 2에 나타내었다. 24 및 48시간 노출 후의 총단백질은 9℃에서 각각 1.6±0.2 및 2.7±0.1 g/dl, NH3는 331.8±13.5 및 428.7±32.3 μg/dl, 삼투질농도는 347.5±5.3 및 344.7±3.4 mmol/kg으로 다른 실험구에 비해 유의한 차이를 보였다. 또한 Na+는 119.0±4.8 및 156.8±0.7 mEq/l, Cl-은 104.2±4.8 및 146.5±0.7 mEq/l로 24 및 48시간 노출 후에 9℃에서 다른 실험구에 비해 증가하였으나, K+는 24시간째에는 12℃에서 1.8±0.2 mEq/l, 48시간 후에는 9℃에서 2.9±0.1 mEq/l로 대조구인 21℃에 비해 유의한 차이를 나타내었다.
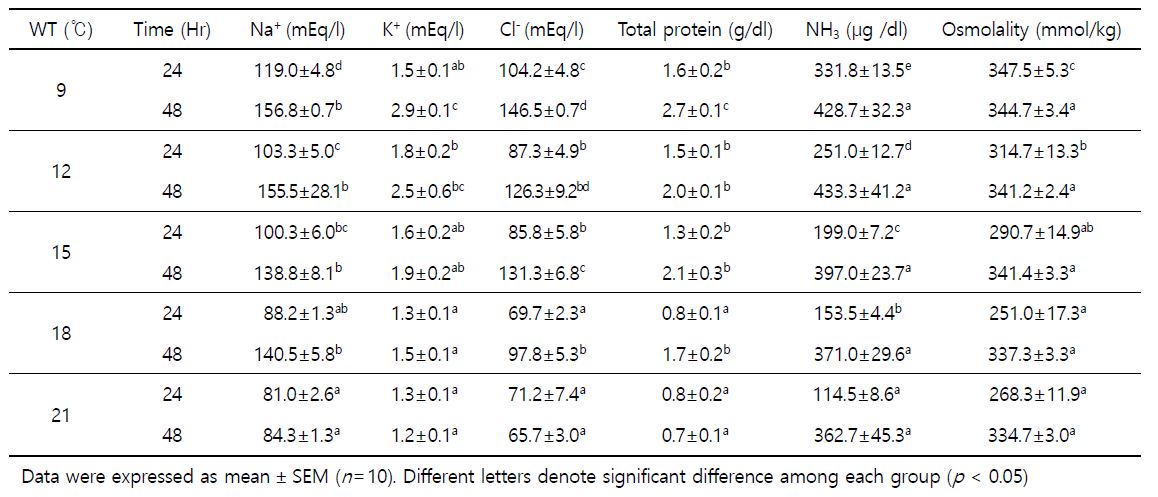
2. Hsp70 mRNA 발현
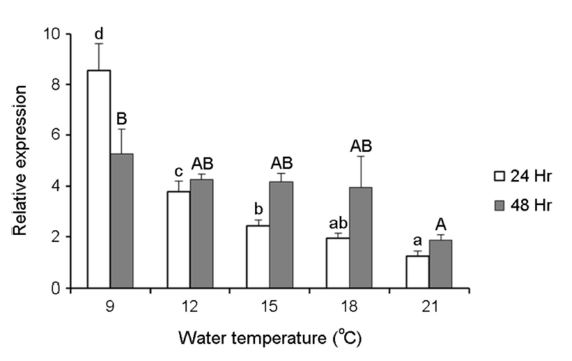
수온변화에 따른 스트레스의 요인이 넙치의 Hsp70 mRNA 발현에 어떠한 영향을 미치는지 알아보기 위해 간 조직으로부터 q-PCR법을 통해 확인하였다. 수온별 Hsp70 mRNA 발현값은 Fig. 5에 나타내었다. 각 수온별 노출 24 및 48시간 노출 후, 9℃가 대조구인 21℃보다 발현량이 유의하게 증가하였다. 24시간 및 48시간 후, 9℃는 각각 8.6±1.0, 5.3±1.0 fold로 21℃의 1.2±0.2, 1.9±0.2 fold보다 높은 발현능을 보였다.
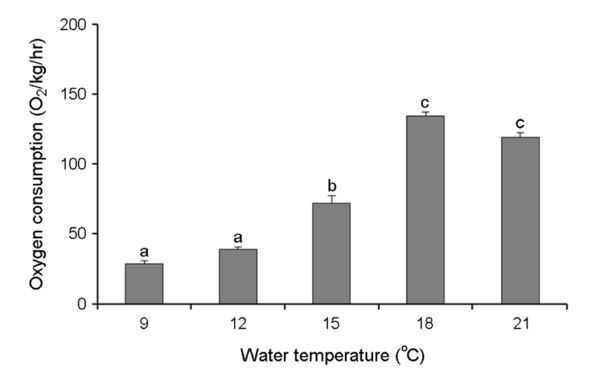
3. 산소소비량 변화
수온별 넙치의 단위 체중당 산소소비량은 Fig. 6에 나타내었다. 9℃에서 28.7±2.3 mg O2/kg/h, 12℃에서 39.3±1.5 mg O2/kg/h, 15℃에서 72.1±5.2 mg O2/kg/h, 18℃에서 134.3±3.0 mg O2/kg/h, 21℃에서 119.1±3.5 mg O2/kg/h로 9 및 12℃에서 21℃에 비해 산소소비량이 유의한 차이를 보였다(Fig. 5). 평균 산소소비량은 수온이 낮아질수록 유의하게 감소하여 9℃에서 가장 감소하였다. 수온이 낮아질수록 산소소비량이 감소하는 것은 수온의 영향에 따른 것으로 나타났다.
고 찰
본 연구에서는 급격한 수온의 변화에 따라 발생하는 스트레스가 넙치에게 어떤 영향을 미치는지 알아보기 위해 혈액학적 분석, Hsp70 mRNA 발현분석 및 산소소비량을 조사하였다. 수온별 모든 실험구의 넙치는 48시간 동안 전량 생존하였다.
각 수온별 넙치의 스트레스 반응에서 대조구인 21℃에 비해 9℃와 12℃에서 다른 실험구보다 유의한 차이를 보이면서, 시작수온인 21℃에서 수온이 급격하게 낮아질수록 적응력이 떨어지는 것을 확인하였다. 혈장 코티졸과 글루코스의 변화는 어류의 스트레스에 있어 주요한 지표이다(Wedemeyer and Yasutake, 1977). 글루코스와 코티졸은 정상적인 어류가 고수온 및 저수온의 조건에서 비례적으로 증가하고, 혈중 함량이 급격하게 증가하는 것이 해산어류인 능성어(Epinephelus septemfasciatus)의 연구에서 보고되어졌으며, 코티졸 농도가 6℃ 및 7℃에서 급격히 증가하고, glucose 또한 5℃ 및 7℃에서 유의하게 증가하였다(Park et al., 2016). 또한, 붉바리(Epinephelus akaara)는 수온자극 2일 및 7일째에 코티졸과 글루코스의 변화량이 유의하게 증가함을 확인하였다(Cho et al., 2015). 이러한 결과는 넙치가 최적수온보다 낮아졌을 때, 급격한 스트레스의 요인으로 인해 혈장 내 코티졸 및 글루코스가 증가하는 결과와 일치한다.
9℃에서 Hb 및 Ht 값의 변화는 24 및 48시간 노출 후에 21℃와 비교했을 때, 유의한 차이를 보였다. 일반적으로, 혈액생리학적 변화는 스트레스 반응에 있어 2차적인 현상으로 나타난다(Barton and Iwama, 1991). 그 중에서도 Hb는 급격한 수온변화에 따른 스트레스를 받았을 때, 산소결합능력을 감소시킴으로써 심각한 영향을 초래한다(Vinodhini and Narayanan, 2009). 이와 더불어 Ht는 혈액의 적혈구 용적비율을 나타내며, 스트레스에 노출되었을 때 생체 내 항상성을 유지시키는데 주요한 역할을 한다(Iwama et al., 1989). 본 연구에서도 마찬가지로, 급격한 수온변화에 따른 스트레스로 인해 넙치의 Ht 및 Hb의 증가를 확인할 수 있었고, 이 결과는 넙치의 생체 내의 산소결합과 수송을 촉진하기 위한 반응에 따라 최적수온의 범위를 벗어난 수온에서 유의한 증가가 나타난 것으로 생각된다.
수온별 넙치의 혈장으로부터 AST 및 ALT, 전해질농도(Na+, K+ and Cl-) 및 삼투질농도, 총단백질 및 NH3를 분석하였다. 첫 번째로, AST 및 ALT는 대부분의 척추동물의 간 세포에 존재하며, 간 기능의 검사지표로 사용된다. 다양한 스트레스 요인에 의해 간 세포가 단기간 및 장기간에 걸쳐 파괴되면 혈액 중에서 이들 효소의 수치가 증가하게 된다(Kang et al., 2007). 특히 어류에서는 주로 간 및 비장세포에 분포하며, 급격한 수온의 변화, 저산소(hypoxia), pH, 암모니아 및 중금속 오염에 의한 스트레스 반응으로 수치가 증가한다(Pan et al., 2003). 또한 Na+, K+, Cl- 및 삼투질 농도에서도 다른 연구와 마찬가지로 급격한 수온변화에 따라 발생된 스트레스를 극복하기 위해 유의적인 차이가 나타난 것으로 보인다. 또한 어류에서 환경변화에 따른 스트레스 요인이 Na+, K+, Cl- 및 삼투질농도의 변화가 해수 및 담수어류의 체내에 일정한 염분을 유지함으로써, 세포 내외의 경사를 유지하기 위하여 이온은 방출하고 물은 흡수하는 저삼투압조절 능력을 가진다(Min et al., 2006). 혈장 내 총단백질과 NH3는 급격한 수온의 변화에 따라 적정수온으로부터 범위를 벗어나 수온이 낮아질수록 다른 실험구에 비해 유의적인 차이를 보였다. 어류의 총단백질량은 건강도, 영양상태 및 질병진단의 지표로 사용되어지고 있고(Nakagawa et al., 1977; Sidduqui, 1977; Byrne et al., 1989), 수온, 수질 및 기타 요인으로부터 발생된 스트레스에 의한 영향이 보고되어진 바 있다(McLeay and Brown, 1979; Ishioka, 1980). NH3는 간 조직 기능의 저하로 인해 혈액 내에서 증가하게 된다(Randall, 2002). 조피볼락(Sebastes schlegeli) (Do et al., 2016), 붉바리(Epinephelus akaara) (Lim et al, 2016), 숭어(Mugil cephalus) 및 나일틸라피아(Oreochromis niloticus) (Chang and Hur, 1999)로부터 수온 및 염분변화에 따른 스트레스 반응과 저산소증에 따른 삼투질농도가 증가하는 것이 보고되었다.
Hsp70 mRNA 발현은 대조구인 21℃에 비해 9℃에서 높았다. 또한 Hsp70은 수온자극, 환경변화 및 오염에 의해 발현되는 스트레스 관련 단백질로 잘 알려져 있고(Beckmann et al., 1990) 특히, 급격한 수온변화에 따라 민감하게 반응하는 단백질로 보고된 바 있으며(Feder and Hofmann, 1999), 다양한 요인에 인한 충격으로부터 생체 내 항상성(homeostasis)을 유지하는데 주요한 역할을 수행한다(Sanders, 1993; Suzue and Young, 1996; Iwama et al., 1998; Ackerman and Iwama, 2001).
수계에 녹아있는 용존산소는 수생생물의 호흡에 반드시 필요한 요소로써, 생리 및 생태학적 조건에 중요한 영향을 미치며, 급격한 수온변화에 따른 대사량 변화를 검증하기 위해 주로 산소소비량 조사를 수행하고 있고(Byun et al., 2008), 이 중 어류의 산소소비량은 수온, 사료섭취량 및 유영행동 등에 따라 차이를 보인다(Fonds and Veldhuis, 1973). 본 연구에서는 수온별 넙치의 산소소비량이 9℃ 및 12℃에서 다른 실험구에 비해 급격히 낮아졌다. 이전의 어류에 관한 산소소비량의 연구에서는, 수온변화에 따른 대왕바리(Epinephelus lanceolatus)로부터 수온이 낮아졌을 때 감소하는 경향을 보였고(Jung et al., 2014), 수온별 pike perch (Stizostedion lucioperca) 자어의 산소소비량 또한 저수온에서 낮아지는 결과를 나타내었다(Zakes and Karpinski, 1999). 따라서, 본 연구의 결과도 마찬가지로 넙치의 적정 수온범위를 벗어나 수온이 낮아질수록 대사량이 떨어지는 것과 비례하여 산소소비량도 9℃ 및 12℃에서 다른 실험구에 비해 유의적인 차이를 보인 것으로 사료된다.
본 연구의 결과들을 바탕으로 수온변화에 따른 넙치의 혈액생리학적 반응, 산소소비량 및 기초적인 분자 및 생리학적 매커니즘을 밝히는데 있어 기초적인 연구보고가 될 것으로 판단되며, 넙치의 장거리 수송 시에 필요한 적정수온 설정에 있어 기초 생물학적 자료로 이용될 수 있을 것으로 사료된다.
- References
-
1. Ackerman PA, Iwama GK. 2001. Physiological and cellular stress responses of juvenile rainbow trout to Vibriosis. Kor J Aquat Anim Health 13: 173-180.


-
2. Barton BA, Iwama GK. 1991. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annu Rev Fish Dis 1: 3-26.
-
3. Beckmann RP, Mizzen LE, Welch WJ. 1990. Interaction of HSP70 with newly synthesized proteins: implications for protein folding and assembly. Sci 248: 850-854.
-
4. Bowden TJ. 2008. Modulation of the immune system of fish by their environment. Fish Shellfish Immunol 25: 373-383.
-
5. Byrne P, Speare DJ, Ferguson HW. 1989. Effects of cationic deter- gent on the gills and blood chemistry of rainbow trout (Salmo gairdneri). Dis Aquat Org 6: 195-196.
-
6. Byun SK, Jeong MH, Lee JH, Lee BI, Ku HD, Park SU, Kim YC, Chang YJ. 2008. Diel rhythm of oxygen consumption of the starry flounder Platichthylys stellatus by water temperature. Kor J Fish Soc 41: 113-118.
-
7. Chang YJ, Hur JW. 1999. Physiological responses of grey mullet (Mugil cepjalus) and Nile tilapia (Oreochromis niloticus) by rapid changes in salinity of rearing water. Kor J Fish Soc 32: 310-316.
-
8. Choi CY, Min BH, Jo PG, Chang YJ. 2007. Molecular cloning of PEPCK and stress response of black porgy (Acanthopagrus schlegeli) to increased temperature in freshwater and seawater. Gen Comp Endocrinol 152: 47-53.
-
9. Cho HC, Kim JE, Kim HB, Baek HJ. 2015. Effects of water tem- perature change on the hematological responses and plasma cortisol levels in growing of red spotted grouper, Epinephelus akaara. Dev Reprod 1: 19-24.
-
10. Cossins AR, Schwarzbaum PJ, Wieser W. 1995. Chapter 6 Effect of temperature on cellular ion regulation and membrane transport systems. Biochem Mol Biol Fish 5: 101-126.
-
11. Do YH, Min BH, Kim YD, Park MS. 2016. Changes on hemato- logical factors and oxygen consumption of Korea rockfish Sebastes schlegeli in high water temperature. JFMSE 28: 738 -745.
-
12. Ellis AE. 2001. Innate host defense mechanisms of fish against viruses and bacteria. Dev Comp Immunol 25: 827-839.
-
13. Feder ME, Hofmann GE. 1999. Heat-shock protein, molecular chap- erones, and the stress response: evolutionary and ecological physiology. Annu Rev Physiol 61: 243-282.
-
14. Fonds M, Veldhuis C. 1973. The oxygen consumption of four Pomatoschistus species (Pisces, Gobiidae) in relation to water temperature, Net J Sea Res 7: 376-386.
-
15. Forsyth RB, Candido EPM, Babich SL, Iwama GK. 1997. Stress pro- tein expression in coho salmon with bacterial kidney disease. J Aquat Anim Health 9: 8-25.
-
16. Iwama GK, McGeer JC, Pawluk MP. 1989. The effects of five fish anaesthetics on acidbase balance, hematocrit, blood gases, cortisol, and adrenaline in rainbow trout. Can J Zool 67: 2065 -2073
-
17. Iwama GK, Thomas PT, Forsyth RB, Vijayan MM. 1998. Heat shock protein expression in fish. Rev Fish Biol 8: 35-36.
-
18. Ishioka H. 1980. Stress reactions in the marine fish. I. Stress re- actions induced by temperature change. Bull Jap Sci Fish 46: 523-532.
-
19. Jung MH, Kim SD, Lim HK, Kim MJ, KIM BS. 2014. Effects of oxygen consumption in the giant grouper Epinephelus lanceolatus by water temperature and photoperiod. Kor J Icthyol 4: 259 -266.
-
20. Kang DY, Kang HW, Kim GH, Jo KC, Kim HC. 2007. Effect of cold shock on the physiological response of the cultured mullet, Mugil haematocheilus in winter. Kor J Fish Soc 40: 226-233.
-
21. Lim SG, Han SB, Lim HK. 2016. Effects of salinity on the growth, survival and stress responses of red spotted grouper Epine- phelus akaara and hybrid grouper E. akaara ♀ × E. lan- ceolatus ♂. Kor J Sci 49: 612-619.
-
22. Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT metheod. Methods 25: 402-408.
-
23. McLeay DJ, Brown DJ. 1979. Stress and chronic effects of untreated and treated bleched keaft pulpmill effluent on the bioche- mistry and stamina of juvenile coho salmon (Oncorhynchus kisutch). Can J Fish Res Board 36: 1049-1059.
-
24. Milani V, Noessner E, Ghose S, Kuppner M, Ahrens B, Scharner A. 2002. Heat shock protein 70: role in antigen presentation and immune stimulation. Inter J Hypertherm 18: 563-575.
-
25. Min BH, Noh GA, Jeong MH, Kang DY, Choi CY, Bang IC, Chang YJ. 2006. Effects of oral administration of thyroid hormone on physiological activity and growth of black porgy reared in fresh water or seawater. Kor J Aquac 19: 149-156.
-
26. Nakagawa H, Kayama, Ikuta K. 1977. Electrophoretic evidence of seasonal variation of carp plasma albumin. J Fac Fish Anim Husb Hiroshima Univ 16: 99-106.
-
27. Nolan DT, Veld RO, Balm PHM, Wendelaar Bonga SE. 1999. Ambi- ent salinity modulates the response of the tilapia, Oreo- chromis mossoambicus (Peters) to net confinement. Aqua 177: 297-309.
-
28. Pan CH, Chien YH, Hunter B. 2003. The resistance to ammonia stress of Penaeus monodon Fabricius juvenile fed diets sup- plemented with astaxanthin. J Exp Mar Biol Ecol 297: 107-118.
-
29. Park JY, Park JM, Hong CK, Kim KM, Cho JK. 2016. Physiological and biochemical of blood on low temperature stress in seven- band grouper, Epinephelus septemfasciatus. Kor J Ichthyol 28: 1-8.
-
30. Perry SF, Reid SD. 1993. β-adrenergic signal transduction in fish: interactive effects of catecholamines and cortisol. Fish Physiol Biochem 11: 195-203.
-
-
32. Schlesinger MJ, Ashburner M, Tissieres A. 1992. Heat shock from bacteria to man. Cold Spring Harbor New York 131-137.
-
33. Siddiqui N. 1977. Seasonal, size and comparative study of plasma proteins of four air breathing freshwater fishes. Proc Indian Acad Sci Sect B 85: 384-390.
-
34. Srivastava P. 2002. Roles of heat-shock proteins in innate and adaptive immunity. Nat Rev Immunol 2: 185-194.
-
35. Suzue K, Young RA. 1996. Adjuvant-free hsp70 fusion protein system elicits humoraland cellular immune responses to HIV-1 p24. J Immunol 156: 873-879.
-
36. Vinodhini R, Narayanan M. 2009. The impact of toxic heavy metal son the hematological parameters in common carp (Cyprinus carpio L.). Iranian J Environ Health Sci Eng 6: 23-28.
-
37. Wedemeyer GA, Yasutake WT. 1977. Clinical methods for the assessment of the effects of environmental stress on fish health. U.S. Fish Wildlife Service Technical 89.
-
38. Wedemeyer GA, Mcleay DJ. 1981. Methods for determining the tolerance of fishes to environmental stressors. In: Stress and Fish. Pickering AD ed Pergamon Press London UK 247-275.
-
39. Yang SJ, JY Lee, JC Jun, JI Myeong, BH Min. 2017. Investigation of suitable temperature and salinity for long distance trans- port of the rockfish Sebastes schlegeli. Kor J Fish Aquat Sci 50: 25-31.
-
40. Zakes Z, Karpinsku A. 1999. Influence of water temperature on oxygen consumption and ammonia excretion of juvenile pike perch, Stizostedion lucioperca (L.) reared in a recirculation system. Aquac Res 2: 109-114.
Copyright (C) 2024. The Korean Society of Marine Life Science